柠檬酸循环
编辑柠檬酸循环(CAC)-也被称为TCA循环(三羧酸循环)或三羧酸循环-是一系列的化学反应所使用的所有需氧微生物通过释放储存的能量的氧化的乙酰-CoA来自碳水化合物、脂肪和蛋白质。此外,循环提供某些氨基酸的前体,以及还原剂NADH,用于许多其他反应。它对许多生化途径的核心重要性表明它是最早的新陈代谢成分之一,可能起源于非生物源。尽管它被标记为“循环”,但代谢物不一定只遵循一个特定的路线;至少已识别出柠檬酸循环的三个部分。
这种代谢途径的名称来源于柠檬酸(一种三羧酸,通常称为柠檬酸盐,因为电离形式在生物pH值下占主导地位),它被消耗,然后通过一系列反应再生以完成循环。该循环消耗乙酸(以乙酰辅酶A的形式)和水,将NAD+还原为NADH,释放二氧化碳。柠檬酸循环产生的NADH被送入氧化磷酸化(电子传输)途径。这两个密切相关的途径的最终结果是营养物质的氧化以ATP的形式产生可用的化学能。
在真核细胞中,柠檬酸循环发生在线粒体基质中。在缺乏线粒体的原核细胞(例如细菌)中,柠檬酸循环反应序列在细胞质中进行,ATP产生的质子梯度穿过细胞表面(质膜)而不是线粒体的内膜。来自TCA循环的含能量化合物的总产率为3个NADH、1个FADH2和1个GTP。
发现
编辑柠檬酸循环的几个组成部分和反应是在1930年代由AlbertSzent-Györgyi的研究确定的,他于1937年获得诺贝尔生理学或医学奖,特别是因为他发现了富马酸,富马酸是循环。他是通过研究鸽子胸肌而发现的。由于这种组织在“Latapie”磨机中分解并在水溶液中释放后仍能很好地保持其氧化能力,因此鸽子的胸肌非常适合用于研究氧化反应。柠檬酸循环本身最终于1937年由汉斯·阿道夫·克雷布斯和威廉·阿瑟·约翰逊,而在谢菲尔德大学,对于其中前者收到的诺贝尔生理学或医学奖于1953年,并为他们的周期有时名为(三羧酸循环)。
概述
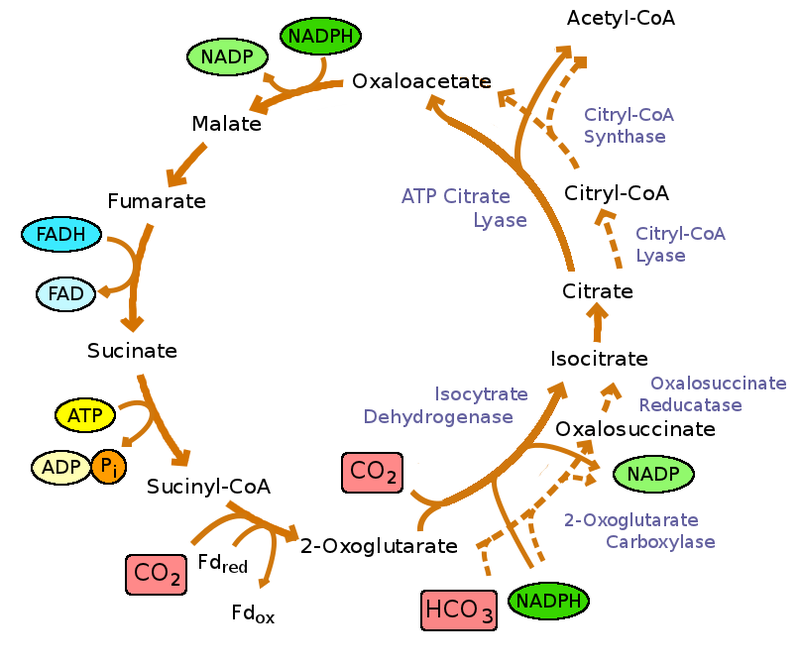
编辑柠檬酸循环是连接碳水化合物、脂肪和蛋白质代谢的关键代谢途径。循环的反应由八种酶进行,它们以乙酰辅酶A的形式将乙酸盐(一个两个碳分子)完全氧化成两个分子,分别为二氧化碳和水。通过糖、脂肪和蛋白质的分解代谢,产生二碳有机产物乙酰辅酶A,进入柠檬酸循环。该循环的反应还将三当量的烟酰胺腺嘌呤二核苷酸(NAD+)转化为三当量的还原NAD+(NADH),将一当量的黄素腺嘌呤二核苷酸(FAD)转化为一当量的FADH2,将鸟苷二磷酸(GDP)和无机磷酸盐(Pi)分别转化为一当量的鸟苷三磷酸盐(GTP)。柠檬酸循环产生的NADH和FADH2反过来被氧化磷酸化途径用于产生富含能量的ATP。
乙酰辅酶A的主要来源之一是糖酵解糖酵解产生的丙酮酸,丙酮酸又被丙酮酸脱氢酶复合物脱羧,产生乙酰辅酶A,根据以下反应方案:
CH3C(=O)C(=O)O-丙酮酸+HSCoA+NAD+→CH3C(=O)SCoA乙酰辅酶A+NADH+CO2
该反应的产物乙酰辅酶A是柠檬酸循环的起点。乙酰辅酶A也可以从脂肪酸的氧化中获得。下面是循环的示意图:
- 的柠檬酸循环开始于一个两碳的转移乙酰基从乙酰-CoA到四碳受体化合物(草酰乙酸盐),以形成六碳化合物(柠檬酸盐)。
- 然后柠檬酸盐经历一系列化学转化,失去两个羧基作为CO2。作为CO2损失的碳来自草酰乙酸,而不是直接来自乙酰辅酶A。在柠檬酸循环的xxx轮之后,由乙酰辅酶A提供的碳成为草酰乙酸碳骨架的一部分。作为CO2的乙酰辅酶A供体碳的损失需要柠檬酸循环的几轮。然而,由于柠檬酸循环在合成代谢中的作用,它们可能不会丢失,因为许多柠檬酸循环中间体也用作其他分子生物合成的前体。
- 通过循环的氧化步骤获得的大部分电子被转移到NAD+,形成NADH。对于进入柠檬酸循环的每个乙酰基,会产生三个NADH分子。柠檬酸循环包括线粒体中的一系列氧化还原反应。
- 此外,来自琥珀酸氧化步骤的电子首先转移到琥珀酸脱氢酶的FAD辅因子,将其还原为FADH2,最终转移到线粒体膜中的泛醌(Q),将其还原为底物泛醇(QH2)复合物III水平的电子转移链。
- 对于在柠檬酸循环中产生的每个NADH和FADH2,在氧化磷酸化中分别产生2.5和1.5个ATP分子。
- 在每个循环结束时,四碳草酰乙酸已经再生,循环继续。
柠檬酸循环的规例
编辑代谢物的变构调节。柠檬酸循环的调节很大程度上取决于产物抑制和底物可用性。如果允许循环不受控制地运行,大量代谢能量可能会浪费在还原型辅酶(如NADH和ATP)的过度生产中。循环的主要最终底物是ADP,它被转化为ATP。减少的ADP量会导致前体NADH的积累,这反过来又会抑制许多酶。NADH是柠檬酸循环中除琥珀酸脱氢酶之外的所有脱氢酶的产物,它抑制丙酮酸脱氢酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶,以及柠檬酸合酶。乙酰辅酶A抑制丙酮酸脱氢酶,而琥珀酰辅酶A抑制α-酮戊二酸脱氢酶和柠檬酸合酶。当用TCA酶进行体外测试时,ATP抑制柠檬酸合酶和α-酮戊二酸脱氢酶;然而,在休息和剧烈运动之间,体内ATP水平的变化不会超过10%。没有已知的变构机制可以解释来自浓度变化小于10%的变构效应物的反应速率的巨大变化。

柠檬酸盐用于反馈抑制,因为它抑制磷酸果糖激酶,这是一种参与糖酵解的酶,可催化果糖1,6-二磷酸酯(丙酮酸的前体)的形成。当柠檬酸盐积累和酶底物减少时,这会阻止恒定的高通量。
受钙调节。钙还用作柠檬酸循环中的调节剂。在细胞活化过程中,线粒体基质中的钙含量可达到数十微摩尔水平。它激活丙酮酸脱氢酶磷酸酶,进而激活丙酮酸脱氢酶复合物。钙还能激活异柠檬酸脱氢酶和α-酮戊二酸脱氢酶。这增加了循环中许多步骤的反应速率,因此增加了整个途径的通量。
转录调控。最近的工作表明,柠檬酸循环中间体与缺氧诱导因子(HIF)的调节之间存在重要联系。HIF在调节氧稳态中发挥作用,是一种靶向血管生成、血管重塑、葡萄糖利用、铁转运和细胞凋亡的转录因子。HIF是组成型合成的,两个关键脯氨酸残基中至少一个的羟基化介导了它们与vonHippelLindauE3泛素连接酶的相互作用复杂,以它们为目标进行快速降解。该反应由脯氨酰4-羟化酶催化。富马酸和琥珀酸已被鉴定为脯氨酰羟化酶的有效抑制剂,从而导致HIF的稳定。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/124482/