细胞表面受体
编辑细胞表面受体是嵌入细胞质膜的受体。它们通过接收(结合)细胞外分子在细胞信号传导中起作用。它们是特殊的整合膜蛋白,允许细胞和细胞外空间之间的通讯。细胞外分子可能是激素、神经递质、细胞因子、生长因子、细胞粘附分子或营养素;它们与受体反应以诱导细胞代谢和活性的变化。在信号转导过程中,配体结合通过细胞膜影响级联化学变化。
细胞表面受体的结构和机制
编辑许多膜受体是跨膜蛋白。有很多种,包括糖蛋白和脂蛋白。数百种不同的受体是已知的,还有更多有待研究。跨膜受体通常根据其三级(三维)结构进行分类。如果三维结构未知,可以根据膜拓扑进行分类。在最简单的受体中,多肽链一次穿过脂质双层,而其他的,如G蛋白偶联受体,交叉多达七次。每个细胞膜可以有几种膜受体,表面分布不同。取决于膜的种类和细胞功能,单个受体也可以不同地分布在不同的膜位置。受体通常聚集在膜表面,而不是均匀分布。
细胞表面受体的机制
已经提出了两种模型来解释跨膜受体的作用机制。
- 二聚化:二聚化模型表明,在配体结合之前,受体以单体形式存在。当激动剂结合发生时,单体结合形成活性二聚体。
- 旋转:与受体细胞外部分结合的配体诱导部分受体跨膜螺旋的旋转(构象变化)。旋转会改变受体的哪些部分暴露在膜的细胞内侧,从而改变受体与细胞内其他蛋白质相互作用的方式。
域
编辑E=细胞外空间P=质膜I=细胞内空间
质膜中的跨膜受体通常可分为三部分。
细胞外结构域
细胞外结构域就在细胞或细胞器的外部。如果多肽链多次穿过双层,则外部结构域包含穿过膜缠绕的环。根据定义,受体的主要功能是识别并响应一种配体。例如,神经递质、激素或原子离子可以各自与细胞外结构域结合,作为与受体偶联的配体。Klotho是一种影响受体以识别配体(FGF23)的酶。
跨膜结构域
两类最丰富的跨膜受体是GPCR和单程跨膜蛋白。在一些受体中,例如烟碱型乙酰胆碱受体,跨膜结构域通过膜或离子通道周围形成蛋白质孔。通过结合适当的配体激活细胞外结构域后,孔变得容易接近离子,然后扩散。在其他受体中,跨膜结构域在结合后会发生构象变化,从而影响细胞内条件。在一些受体中,例如7TM超家族的成员,跨膜结构域包括一个配体结合口袋。
细胞内结构域
受体的细胞内(或细胞质)结构域与细胞或细胞器内部相互作用,传递信号。这种交互有两种基本途径:
信号转导
编辑通过膜受体的信号转导过程涉及配体与膜受体结合的外部反应和触发细胞内反应的内部反应。
通过膜受体进行信号转导需要四个部分:
- 细胞外信号分子:细胞外信号分子由一个细胞产生,至少能够传播到相邻细胞。
- 受体蛋白:细胞必须具有细胞表面受体蛋白,这些受体蛋白与信号分子结合并向内传递到细胞中。
- 细胞内信号蛋白:它们将信号传递给细胞的细胞器。信号分子与受体蛋白的结合将激活启动信号级联的细胞内信号蛋白。
- 靶蛋白:当信号通路激活并改变细胞行为时,靶蛋白的构象或其他特性会发生改变。
乙酰胆碱受体的三种构象状态(点击放大)
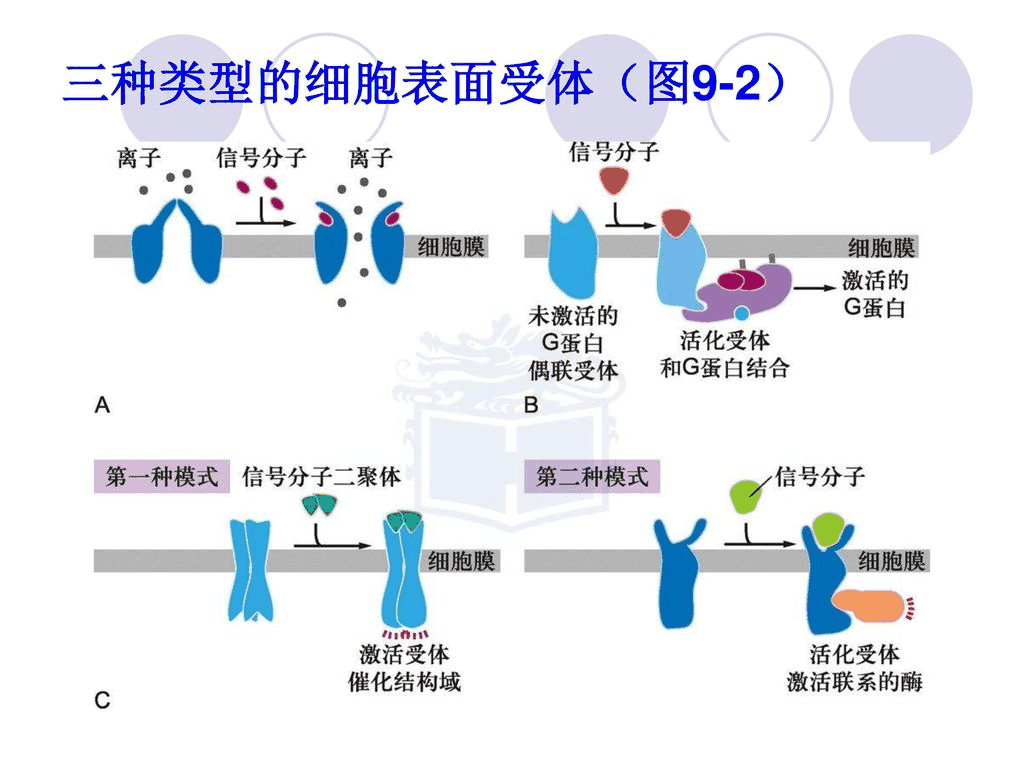
膜受体主要按结构和功能分为3类:离子通道连接受体;酶联受体;和G蛋白偶联受体。
- 离子通道连接受体具有阴离子和阳离子的离子通道,并构成一个大家族的多通道跨膜蛋白。它们参与通常在神经元等电活性细胞中发现的快速信号传导事件。它们也被称为配体门控离子通道。离子通道的打开和关闭由神经递质控制。
- 酶联受体要么是酶本身,要么是直接激活相关酶。这些通常是单程跨膜受体,受体的酶成分保持在细胞内。大多数酶联受体是蛋白激酶或与蛋白激酶相关。
- G蛋白偶联受体是具有七个跨膜螺旋的完整膜蛋白。这些受体在激动剂结合后激活G蛋白,而G蛋白介导受体对细胞内信号通路的影响。
离子通道相关受体
在神经元中的信号转导过程中,神经递质与受体结合并改变蛋白质的构象。这打开了离子通道,允许细胞外离子进入细胞。质膜的离子通透性被改变,这将细胞外化学信号转化为细胞内电信号,从而改变细胞的兴奋性。
乙酰胆碱受体是与阳离子通道相连的受体。该蛋白质由四个亚基组成:α(α)、β(β)、γ(γ)和δ(δ)亚基。有两个α亚基,一个乙酰胆碱每个绑定站点。该受体可以以三种构象存在。封闭和空置状态是天然蛋白质构象。由于两个乙酰胆碱分子都与α亚基上的结合位点结合,因此受体的构象发生改变,门打开,允许许多离子和小分子进入。但是,这种打开和占用的状态只持续了很短的时间,然后门就关闭了,变成了关闭和占用的状态。两个乙酰胆碱分子将很快从受体上解离,使其恢复到天然的封闭和空置状态。
酶联受体
截至2009年,已知有6种酶联受体:受体酪氨酸激酶;酪氨酸激酶相关受体;受体样酪氨酸磷酸酶;受体丝氨酸/苏氨酸激酶;受体鸟苷酸环化酶和组氨酸激酶相关受体。受体酪氨酸激酶具有xxx的种群和最广泛的应用。这些分子中的大多数是生长因子的受体,例如表皮生长因子(EGF)、血小板衍生生长因子(PDGF)、成纤维细胞生长因子(FGF)、肝细胞生长因子(HGF)、神经生长因子(NGF)和胰岛素等激素。大多数这些受体在与其配体结合后会二聚化,以激活进一步的信号转导。例如,在表皮生长因子(EGF)受体与其配体EGF结合后,这两个受体发生二聚化,然后每个受体分子的酶部分中的酪氨酸残基发生磷酸化。这将激活酪氨酸激酶并催化进一步的细胞内反应。
G蛋白偶联受体
G蛋白偶联受体包括一个大的跨膜受体蛋白家族。它们只存在于真核生物中。结合和激活这些受体的配体包括:光敏化合物、气味、信息素、激素和神经递质。它们的大小从小分子到肽和大蛋白质不等。G蛋白偶联受体与许多疾病有关,因此是许多现代药物的靶点。
涉及G蛋白偶联受体的主要信号转导途径有两条:cAMP信号传导途径和磷脂酰肌醇信号传导途径。两者都是通过G蛋白激活介导的。G蛋白是一种三聚体蛋白,具有三个亚基,分别称为α、β和γ。响应受体激活,α亚基释放结合的鸟苷二磷酸(GDP),其被三磷酸鸟苷(GTP)取代,从而激活α亚基,然后从β和γ亚基解离。活化的α亚基可以进一步影响细胞内信号蛋白或直接靶向功能蛋白。
膜受体相关疾病
编辑如果膜受体变性或缺陷,信号转导就会受到阻碍并引起疾病。有些疾病是由膜受体功能障碍引起的。这是由于编码和调节受体蛋白的基因发生变化导致受体缺乏或降解。膜受体TM4SF5影响肝细胞和肝癌的迁移。此外,皮质NMDA受体影响膜流动性,并在阿尔茨海默病中发生改变。当细胞被无包膜病毒感染时,病毒首先与特定的膜受体结合,然后将自身或亚病毒成分传递到细胞膜的细胞质侧。如果是脊髓灰质炎病毒,在体外已知与受体的相互作用引起构象重排,从而释放出称为VP4的病毒粒子蛋白。VP4的N末端是肉豆蔻基化的,因此具有疏水性【肉豆蔻酸=CH3(CH2)12COOH】。有人提出,受体结合诱导的构象变化导致肉豆蔻酸附着在VP4上并形成RNA通道。
基于结构的药物设计
编辑通过X射线晶体学和核磁共振波谱等方法,靶分子的3D结构信息急剧增加,配体的结构信息也急剧增加。这推动了基于结构的药物设计的快速发展.其中一些新药靶向膜受体。当前基于结构的药物设计方法可分为两类。xxx类是关于确定给定受体的配体。这通常通过数据库查询、生物物理模拟和化学库的构建来完成。在每种情况下,都会筛选大量潜在的配体分子以找到适合受体结合口袋的那些。这种方法通常被称为基于配体的药物设计。搜索数据库的主要优势在于它可以节省时间和精力来获得新的有效化合物。基于结构的药物设计的另一种方法是关于组合映射配体,这被称为基于受体的药物设计。在这种情况下,通过以逐步方式组装小片段,在结合口袋的约束下设计配体分子。这些碎片可以是原子或分子。这种方法的主要优点是可以发现新的结构。

其他例子
编辑- 肾上腺素受体
- 嗅觉受体
- 受体酪氨酸激酶
- 表皮生长因子受体
- 胰岛素受体
- 成纤维细胞生长因子受体
- 高亲和力神经营养因子受体
- 肝配蛋白受体
- 整合素
- 低亲和力神经生长因子受体
- NMDA受体
- 几种免疫受体
- Toll样受体
- T细胞受体
- CD28
- SCIMP蛋白
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/133931/