
DNA连接酶
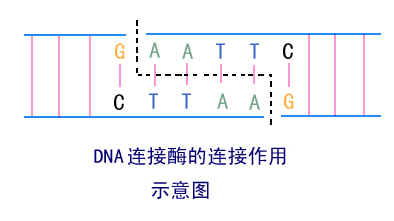
编辑DNA 连接酶是一种特殊类型的酶,一种连接酶 (EC 6.5.1.1),它通过催化磷酸二酯键的形成来促进 DNA 链的连接。 它在修复生物体双链 DNA 中的单链断裂方面发挥作用,但某些形式(如 DNA 连接酶 IV)可能专门修复双链断裂(即 DNA 两条互补链的断裂)。 单链断裂由 DNA 连接酶修复,使用双螺旋的互补链作为模板,DNA 连接酶产生最终的磷酸二酯键以完全修复 DNA。
DNA 连接酶用于 DNA 修复和 DNA 复制(参见哺乳动物连接酶)。 此外,DNA 连接酶在分子生物学实验室中广泛用于重组 DNA 实验(参见研究应用)。 纯化的 DNA 连接酶用于基因克隆,将 DNA 分子连接在一起形成重组 DNA。
酶促机制
编辑DNA连接酶的机制是在一个核苷酸(受体)的3′羟基末端与另一个核苷酸(供体)的5’磷酸末端之间形成两个共价磷酸二酯键。 每个形成的磷酸二酯键消耗两个 ATP 分子。 连接酶反应需要 AMP,该反应分四个步骤进行:
- 活性位点的重组,如DNA片段中的缺口或冈崎片段等。
- 酶活性中心赖氨酸残基的腺苷酸化(AMP 加成),释放出焦磷酸盐;
- 将 AMP 转移到所谓供体的 5′ 磷酸,形成焦磷酸键;
- 在供体的 5′ 磷酸和受体的 3′ 羟基之间形成磷酸二酯键。
连接酶也适用于平端,但需要更高的酶浓度和不同的反应条件。
类型
编辑E. 大肠杆菌
编辑大肠杆菌 DNA 连接酶由 lig 基因编码。 大肠杆菌以及大多数原核生物中的 DNA 连接酶使用通过裂解烟酰胺腺嘌呤二核苷酸 (NAD) 获得的能量来产生磷酸二酯键。 它不连接平端 DNA,除非在聚乙二醇分子拥挤的条件下,并且不能有效地将 RNA 连接到 DNA。
适当浓度的 DNA 聚合酶可增强大肠杆菌 DNA 连接酶的活性。 只有当 DNA 聚合酶 1 的浓度远低于要连接的 DNA 片段时,增强才会起作用。 当 Pol I DNA 聚合酶的浓度较高时,它对大肠杆菌 DNA 连接酶有不利影响
T4
编辑来自噬菌体 T4(一种感染大肠杆菌的噬菌体)的 DNA 连接酶。 T4 连接酶是实验室研究中最常用的连接酶。 它可以连接 DNA、寡核苷酸以及 RNA 和 RNA-DNA 杂交体的粘性末端或平末端,但不能连接单链核酸。 它还可以比大肠杆菌 DNA 连接酶更有效地连接平端 DNA。 与大肠杆菌 DNA 连接酶不同,T4 DNA 连接酶不能利用 NAD,它绝对需要 ATP 作为辅助因子。 已经进行了一些工程来提高 T4 DNA 连接酶的体外活性; 例如,一种成功的方法测试了与几种替代 DNA 结合蛋白融合的 T4 DNA 连接酶,发现以 p50 或 NF-kB 作为融合伙伴的构建体在用于克隆目的的平端连接中的活性比野生的高 160% 以上 T4 型 DNA 连接酶。 将片段插入质粒载体的典型反应将使用约 0.01(粘性末端)至 1(平末端)单位的连接酶。 T4 DNA 连接酶的最佳孵育温度为 16 °C。
噬菌体 T4 连接酶突变体对紫外线照射和烷化剂甲磺酸甲酯的敏感性增加,表明 DNA 连接酶用于修复由这些试剂引起的 DNA 损伤。
哺乳动物
编辑在哺乳动物中,有四种特定类型的连接酶。
- DNA 连接酶 I:在核糖核酸酶 H 从冈崎片段中去除 RNA 引物后,连接滞后链的新生 DNA。
- DNA 连接酶 III:与 DNA 修复蛋白 XRCC1 形成复合物,有助于在核苷酸切除修复和重组片段过程中密封 DNA。 在所有已知的哺乳动物 DNA 连接酶中,只有 Lig III 被发现存在于线粒体中。
- DNA连接酶IV:与XRCC4复合。 它催化非同源末端连接 DNA 双链断裂修复途径的最后一步。 它也是 V(D)J 重组所必需的,V(D)J 重组是在免疫系统发育过程中产生免疫球蛋白和 T 细胞受体位点多样性的过程。
来自真核生物和一些微生物的 DNA 连接酶使用三磷酸腺苷 (ATP) 而不是 NAD。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229289/