
神经干细胞
编辑显微解剖学的解剖学术语 一些神经祖细胞干细胞持续存在于成年脊椎动物大脑中高度受限的区域,并在整个生命过程中继续产生神经元。 中枢神经系统大小的差异是物种之间最重要的区别之一,因此调节神经干细胞区室大小的基因突变是脊椎动物进化的最重要驱动因素之一。
干细胞的特征在于它们能够分化成多种细胞类型。 它们经历对称或不对称的细胞分裂成两个子细胞。 在对称细胞分裂中,两个子细胞也是干细胞。 在不对称分裂中,一个干细胞产生一个干细胞和一个特化细胞。 NSC 主要分化为神经元、星形胶质细胞和少突胶质细胞。
大脑位置
编辑在成年哺乳动物的大脑中,海马齿状回的颗粒下区、侧脑室周围的脑室下区和下丘脑(恰好位于背侧α1、α2区和下丘脑增殖区,位于相邻的正中隆起处)有 据报道含有神经干细胞。
发展
编辑体内来源
有两种基本类型的干细胞:成体干细胞,其分化能力有限;胚胎干细胞 (ESC),具有多能性并具有分化成任何细胞类型的能力。
神经干细胞比胚胎干细胞更专业,因为它们只产生放射状胶质细胞,这些细胞产生神经元和中枢神经系统 (CNS) 的胶质细胞。 在脊椎动物的胚胎发育过程中,神经干细胞转变为放射状神经胶质细胞 (RGC),也称为放射状神经胶质祖细胞 (RGP),并位于称为心室区 (VZ) 的瞬变区中。 神经元在胚胎发育的特定时期通过神经发生过程由 (RGPs) 大量产生,并在成人大脑的受限区域中继续在成年生活中产生。 成人神经干细胞在成人脑室下区 (SVZ)、胚胎生发神经上皮的残余物以及海马齿状回内分化成新的神经元。
体外来源
成年神经干细胞于 1990 年代初首次从小鼠纹状体中分离出来。 它们在体外培养时能够形成多能神经球。 神经球可以产生自我更新和增殖的特化细胞。 这些神经球可以分化形成特定的神经元、神经胶质细胞和少突胶质细胞。 在以前的研究中,培养的神经球已被移植到免疫缺陷新生小鼠的大脑中,并显示出植入、增殖和神经分化。
交流与迁移
NSC 通过来自微环境或干细胞生态位的外源性信号被刺激开始分化。 一些神经细胞沿着延髓迁移流从 SVZ 迁移,该迁移流在受到刺激时包含具有室管膜细胞和星形胶质细胞的骨髓样结构。 室管膜细胞和星形胶质细胞形成胶质管,供迁移的成神经细胞使用。 管中的星形胶质细胞为迁移细胞提供支持,并隔离周围细胞释放的电信号和化学信号。 星形胶质细胞是快速细胞扩增的主要前体。 成神经细胞形成紧密链并向细胞损伤的特定部位迁移以修复或替换神经细胞。 一个例子是成神经细胞向嗅球迁移以分化成具有径向迁移模式而不是切向迁移模式的球周或颗粒神经元。
老化
神经干细胞的增殖随着年龄的增长而下降。 已采取各种方法来抵消这种与年龄相关的衰退。 由于 FOX 蛋白调节神经干细胞稳态,FOX 蛋白已被用于通过抑制 Wnt 信号来保护神经干细胞。
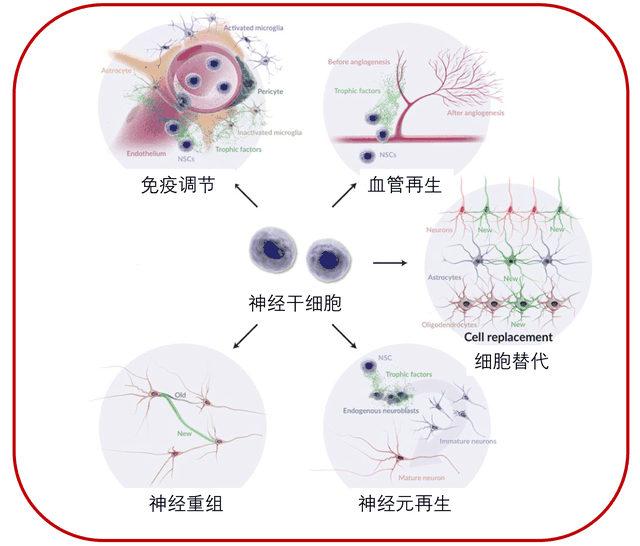
函数
编辑表皮生长因子 (EGF) 和成纤维细胞生长因子 (FGF) 是促进神经祖细胞和干细胞体外生长的有丝分裂原,尽管神经祖细胞和干细胞群合成的其他因子也是最佳生长所必需的。 据推测,成人大脑中的神经发生起源于神经干细胞。 成人大脑中神经干细胞的起源和身份仍有待确定。
分化过程中
最广泛接受的成人 NSC 模型是放射状神经胶质原纤维酸性蛋白阳性细胞。 静止干细胞是 B 型,由于 SPe 提供的可再生组织,它们能够保持静止状态。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/250269/