
简介
编辑化学渗透是离子沿着其电化学梯度穿过半透膜结合结构的运动。一个重要的例子是在细胞呼吸或光合作用过程中氢离子 (H+) 跨膜移动形成三磷酸腺苷 (ATP)。
氢离子或质子将从质子浓度高的区域扩散到质子浓度较低的区域,并且可以利用跨膜质子的电化学浓度梯度来制造 ATP。这个过程与渗透作用有关,渗透作用是水穿过选择性膜的运动,这就是它被称为化学渗透作用的原因。
ATP合酶是通过化学渗透作用产生ATP的酶。它允许质子穿过膜并利用自由能差使二磷酸腺苷 (ADP) 磷酸化,从而生成 ATP。 通过化学渗透作用产生 ATP 在线粒体和叶绿体中,以及大多数细菌和古细菌中。 例如,在光合作用期间的叶绿体中,电子传输链将基质(流体)中的 H+ 离子(质子)通过类囊体膜泵送到类囊体空间。 当质子通过 ATP 合酶移动时,储存的能量用于光磷酸化 ADP,从而产生 ATP。
化学渗透理论
编辑Peter D. Mitchell 于 1961 年提出了化学渗透假说。该理论表明,呼吸细胞中的大多数三磷酸腺苷 (ATP) 合成来自线粒体内膜的电化学梯度,利用分解形成的 NADH 和 FADH2 的能量富含能量的分子,例如葡萄糖。
葡萄糖等分子被代谢产生乙酰辅酶 A 作为一种能量相当丰富的中间体。线粒体基质中乙酰辅酶 A (acetyl-CoA) 的氧化与烟酰胺腺嘌呤二核苷酸 (NAD) 和黄素腺嘌呤二核苷酸 (FAD) 等载体分子的还原偶联。载体将电子传递至电子传递链 ( ETC)在内线粒体膜中,然后将它们传递给 ETC 中的其他蛋白质。 ETC 中的末端受体氧气的能量用于将质子从基质泵入膜间隙,以跨膜电化学梯度的形式储存能量。 质子通过 ATP 合成酶移回内膜。 质子通过 ATP 合酶流回线粒体基质,为 ADP 与无机磷酸盐结合形成 ATP 提供足够的能量。
这在当时是一个激进的提议,并没有被广泛接受。 流行的观点是,电子转移的能量被存储为稳定的高电位中间体,这是一个化学上更保守的概念。旧范式的问题是从未发现高能中间体,电子转移链复合物泵浦质子的证据变得太大而无法忽视。最终,大量证据开始支持化学渗透假说,彼得·米切尔于 1978 年获得诺贝尔化学奖。
化学渗透偶联对于线粒体、叶绿体以及许多细菌和古细菌中 ATP 的产生很重要。
质子动力
编辑离子跨膜的运动取决于两个因素的组合:
- 由浓度梯度引起的扩散力 – 所有粒子都倾向于从较高浓度扩散到较低浓度。
- 由电势梯度引起的静电力 – 像质子 H+ 这样的阳离子倾向于向下扩散电势,从膜的正 (P) 侧到负 (N) 侧。 阴离子自发地向相反方向扩散。
这两个梯度一起可以表示为电化学梯度。
然而,生物膜的脂质双层是离子的屏障。 这就是为什么能量可以作为跨膜的这两个梯度的组合存储的原因。 只有特殊的膜蛋白,如离子通道,有时才能让离子穿过膜。
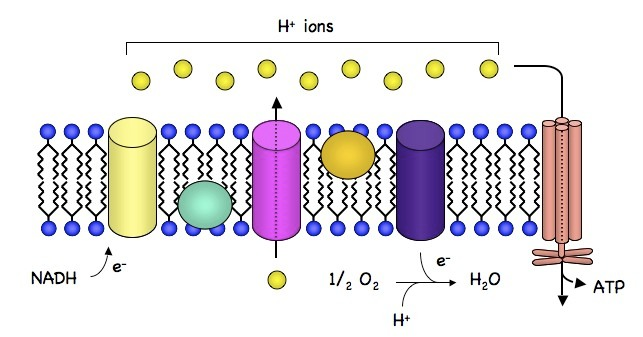
在化学渗透理论中,跨膜 ATP 合酶非常重要。 它们将质子自发流过的能量转化为 ATP 键的化学能。
因此,研究人员创造了质子动力 (PMF) 一词,它源自前面提到的电化学梯度。 它可以描述为存储的势能(化学渗透势)的量度,作为跨膜的质子和电压(电势)梯度的组合。 电梯度是跨膜电荷分离的结果(当质子 H+ 在没有抗衡离子(例如氯离子 Cl−)的情况下移动时)。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228844/