
DNA超螺旋
编辑DNA超螺旋是指特定DNA链中的扭曲量,它决定了其上的应变量。 给定的链可能是正超螺旋或负超螺旋(或多或少紧密缠绕)。 一条链的超螺旋量会影响许多生物过程,例如压缩 DNA 和调节对遗传密码的访问(这会强烈影响 DNA 代谢和可能的基因表达)。 某些酶,如拓扑异构酶,会改变 DNA 超旋的数量,以促进 DNA 复制和转录等功能。 给定链中的超螺旋量由数学公式描述,该公式将其与称为松弛 B 型 DNA 的参考状态进行比较。
概览
编辑在 B-DNA 的松弛双螺旋片段中,两条链每 10.4–10.5 个碱基对序列围绕螺旋轴扭转一次。 像某些酶一样,增加或减少扭曲会施加压力。 如果一个 DNA 片段在扭曲应变下通过连接其两端而闭合成一个圆圈,然后允许其自由移动,它就会呈现出不同的形状,例如数字 8。 这种形状被称为超螺旋。 (在描述 DNA 拓扑结构时经常使用名词形式超螺旋。)
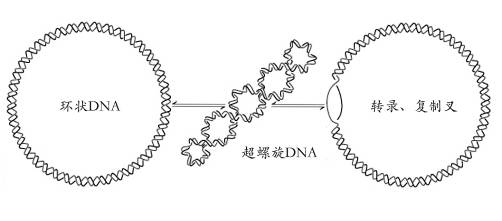
大多数生物体的 DNA 通常是负超螺旋的。 当它被复制或转录时,它会暂时变成正超螺旋。 如果不及时放松,这些过程将被抑制(调节)。 最简单的超螺旋形状是数字 8; 环状 DNA 链呈现这种形状以适应或多或少的螺旋扭曲。 数字 8 的两个叶瓣将相对于彼此顺时针或逆时针旋转,具体取决于螺旋线是过度缠绕还是缠绕不足。 对于每个额外的螺旋扭曲,叶片将显示围绕其轴的更多旋转。
环状 DNA 的局部扭曲,例如上面八字形叶的旋转,被称为扭动。 上面的例子说明了扭曲和扭动是可以相互转换的。 超螺旋可以用扭曲和扭动的总和在数学上表示。 扭曲是 DNA 中的螺旋圈数,而扭曲是双螺旋自身交叉的次数(这些是超螺旋)。 额外的螺旋扭曲是正的并导致正超螺旋,而减法扭曲导致负超螺旋。 许多拓扑异构酶感知超螺旋,并在它们改变 DNA 拓扑结构时产生或消散它。
部分原因是染色体可能非常大,中间的片段可能就像它们的末端被锚定一样。 因此,它们可能无法将多余的扭曲分配给染色体的其余部分,或者无法吸收扭曲以从卷绕中恢复——换句话说,这些片段可能会变成超螺旋。 作为对超螺旋的响应,它们会发生一定程度的扭动,就好像它们的末端是连在一起的一样。
超螺旋 DNA 形成两种结构; 一个 plectoneme 或一个环形线圈,或两者的组合。 一个负超螺旋的 DNA 分子将产生一个单起点左手螺旋,环形,或一个带有末端环的双起点右手螺旋,即 plectoneme。 Plectonemes 通常在自然界中更常见,这是大多数细菌质粒的形状。 对于较大的分子,通常会形成混合结构——环形上的一个环可以延伸到一个 plectoneme 中。 如果环形上的所有环都延伸,那么它就成为 pleconemic 结构中的一个分支点。 DNA 超旋对于所有细胞内的 DNA 包装都很重要,并且似乎也在基因表达中发挥作用。
嵌入诱导的 DNA 超螺旋
编辑基于嵌入分子的特性,即在与 DNA 结合和 DNA 碱基对解旋时发出荧光,2016 年,引入了一种单分子技术来直接可视化沿着超螺旋 DNA 的单个 plectonemes,这将进一步研究相互作用 具有超螺旋 DNA 的 DNA 加工蛋白质。 在该研究中,Sytox Orange(一种嵌入染料)被用于诱导表面拴系 DNA 分子的超螺旋。
使用该测定,发现 DNA 序列编码了 plectonemic 超螺旋的位置。 此外,发现 DNA 超螺旋在原核生物的转录起始位点富集。
函数
编辑基因组包装
编辑DNA超旋对于所有细胞内的DNA包装都很重要。 由于 DNA 的长度可能是细胞的数千倍,因此将这种遗传物质包装到细胞或细胞核(在真核生物中)是一项艰巨的任务。 DNA 的超螺旋减少了空间并允许 DNA 被包装。 在原核生物中,由于环状染色体和相对少量的遗传物质,多核超螺旋占主导地位。 在真核生物中,DNA 超螺旋存在于多个水平的全基因超螺旋和螺线管超螺旋中,螺线管超螺旋被证明在比较中最有效。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229281/