
位点特殊性重组
编辑位点特异性重组,也称为保守的位点特异性重组,是一种遗传重组,其中DNA链交换发生在至少具有一定序列同源性的片段之间。 称为位点特异性重组酶 (SSR) 的酶通过识别和结合短的特定 DNA 序列(位点)来执行 DNA 片段的重排,在这些位点上它们切割 DNA 主链,交换所涉及的两个 DNA 螺旋,并重新加入 DNA 链。 在某些情况下,重组酶和重组位点的存在足以使反应进行; 在其他系统中,需要许多辅助蛋白和/或辅助位点。 许多不同的基因组修饰策略,在这些重组酶介导的盒式交换 (RMCE) 中,是一种将转录单元定向引入预定基因组位点的高级方法,都依赖于 SSR。
位点特殊重组系统具有高度特异性、快速和高效,即使面对复杂的真核基因组。 它们自然地用于各种细胞过程,包括细菌基因组复制、分化和发病机制,以及移动遗传元件的移动。 出于同样的原因,它们为基因工程工具的开发提供了潜在的基础。
重组位点的长度通常在 30 到 200 个核苷酸之间,由两个具有部分反向重复对称性的基序组成,重组酶与其结合,并且位于发生重组的中心交叉序列的侧翼。 发生重组的位点对通常是相同的,但也有例外(例如 λ 整合酶的 attP 和 attB)。
分类:酪氨酸重组酶与丝氨酸重组酶
编辑基于氨基酸序列同源性和机制相关性,大多数位点特异性重组酶分为两个家族之一:酪氨酸 (Tyr) 重组酶家族或丝氨酸 (Ser) 重组酶家族。 这些名称源于每类重组酶中存在的保守亲核氨基酸残基,这些重组酶用于攻击 DNA 并在链交换过程中与其共价连接。 最早发现的丝氨酸重组酶家族成员被称为分解酶或 DNA 转化酶,而酪氨酸重组酶的创始成员 λ 噬菌体整合酶(使用 attP/B 识别位点)不同于现在众所周知的酶,例如 Cre( 来自 P1 噬菌体)和 FLP(来自酵母酿酒酵母)。 著名的丝氨酸重组酶包括诸如 gamma-delta 分解酶(来自 Tn1000 转座子)、Tn3 分解酶(来自 Tn3 转座子)和 φC31 整合酶(来自 φC31 噬菌体)等酶。
虽然两个重组酶家族的个别成员可以进行具有相同实际结果的反应,但这些家族彼此无关,具有不同的蛋白质结构和反应机制。 与酪氨酸重组酶不同,丝氨酸重组酶是高度模块化的,这一点首先由生化研究暗示,随后由晶体结构显示。 当试图重新设计重组酶蛋白质作为基因操作的工具时,这些蛋白质结构的知识可能会被证明是有用的。
机制
编辑两个 DNA 位点之间的重组始于重组酶对这些位点的识别和结合——两个独立的双链 DNA 分子各有一个位点,或同一分子的至少两个远距离片段。 随后是突触,即将这些位点聚集在一起形成突触复合体。 正是在这个突触复合体中,链交换发生了,因为 DNA 通过受控的酯交换反应被切割和重新结合。 在链交换过程中,每个双链 DNA 分子在识别位点交叉区域内的固定点被切割,释放脱氧核糖羟基,而重组酶与 DNA 骨架磷酸盐形成瞬时共价键。 亲核丝氨酸或酪氨酸残基的羟基之间的磷酸二酯键保存了切割 DNA 时消耗的能量。
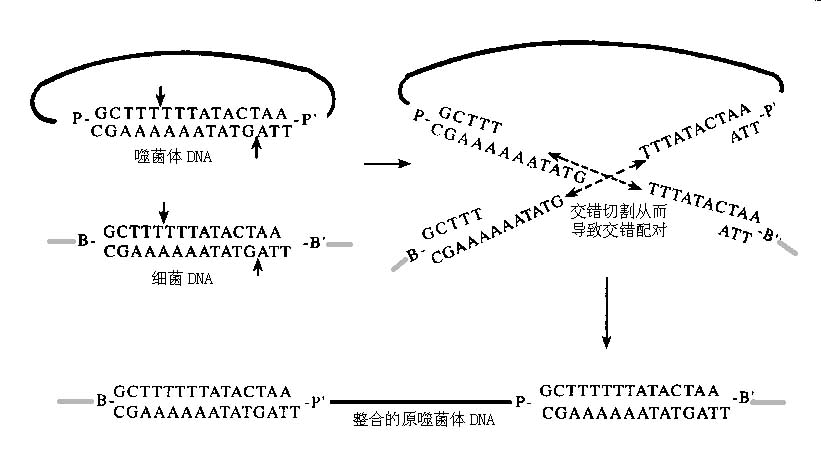
存储在该键中的能量随后用于将 DNA 重新连接到另一个 DNA 分子上相应的脱氧核糖羟基。 因此,整个反应的进行不需要外部富含能量的辅助因子,例如 ATP。
尽管酪氨酸和丝氨酸重组酶的基本化学反应相同,但它们之间存在一些差异。 酪氨酸重组酶,如 Cre 或 FLP,每次在 6-8bp 交错的点切割一条 DNA 链,将链的 3′ 端连接到酪氨酸亲核试剂的羟基。 然后,链交换通过类似于 Holliday 连接的交叉链中间体进行,其中仅交换了一对链。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/250181/