
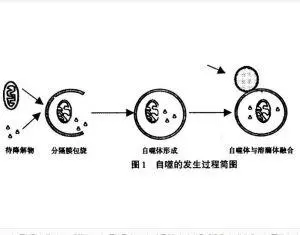
自噬体
编辑自噬体是具有双层膜的球形结构。 它是巨自噬的关键结构,巨自噬是细胞质内容物(例如,异常的细胞内蛋白质、过量或受损的细胞器、入侵的微生物)的细胞内降解系统。 形成后,自噬体将细胞质成分输送到溶酶体。 自噬体的外膜与溶酶体融合形成自噬溶酶体。 溶酶体的水解酶降解自噬体输送的内容物及其内膜。
自噬体的形成受从酵母到高等真核生物都非常保守的基因调控。 这些基因的命名因论文而异,但近年来已被简化。 以前称为 APG、AUT、CVT、GSA、PAZ 和 PDD 的基因家族现在统一为 ATG(AuTophaGy 相关)家族。
自噬体的大小因哺乳动物和酵母而异。 酵母自噬体约为 500-900 nm,而哺乳动物自噬体更大 (500-1500 nm)。 在一些细胞实例中,如胚胎干细胞、胚胎成纤维细胞和肝细胞,自噬体在光学显微镜下可见,并且可以被视为环形结构。
自我形成
编辑自噬体在内质网上形成 omegasome 的初始步骤,随后是称为吞噬泡的结构的伸长。
自噬体的形成由 Atg 基因通过 Atg12-Atg5 和 LC3 复合物控制。 Atg12-Atg5 的偶联物也与 Atg16 相互作用形成更大的复合物。 Atg12 对 Atg5 的修饰对于初始膜的伸长至关重要。
球形结构形成后,ATG12-ATG5:ATG16L1 复合体从自噬体上解离出来。 LC3 被 ATG4 蛋白酶切割以产生胞质 LC3。 自噬体与其靶膜的末端融合需要 LC3 切割。 LC3 通常用作免疫细胞化学中自噬体的标志物,因为它是囊泡的重要组成部分,并且在融合前的最后一刻保持结合。 首先,自噬体与核内体或核内体衍生的囊泡融合。 然后将这些结构称为两性体或中间自噬泡。 尽管如此,这些结构包含内吞标记,甚至是小的溶酶体蛋白,如组织蛋白酶 D。
这个过程在酵母中是相似的,但是基因名称不同。 例如,哺乳动物中的 LC3 是酵母中的 Atg8,而自噬体是由前自噬体结构 (PAS) 生成的,它与哺乳动物细胞中的前体结构不同。 酵母中的前自噬体结构被描述为位于液泡附近的复合物。 然而,这种本地化的意义尚不清楚。 成熟的酵母自噬体直接与液泡或溶酶体融合,不像哺乳动物那样形成两性体。
在酵母自噬体成熟过程中,还有其他已知参与者,如 Atg1、Atg13 和 Atg17。
Atg1 是一种在诱导自噬后上调的激酶。 Atg13 调节 Atg1,它们一起形成一个名为 Atg13:Atg1 的复合体,它接收来自营养感应大师 Tor 的信号。 Atg1 在自噬体形成的后期也很重要。
在神经元中的功能
编辑在神经元中,自噬体在神经突尖端产生,并在它们沿着轴突向细胞体移动时成熟(酸化)。 如果亨廷顿蛋白或其相互作用的伙伴 HAP1(与神经元中的自噬体共定位)被耗尽,这种轴突运输就会被破坏。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228809/