生殖细胞系
编辑在生物学和遗传学中,种系是将遗传物质传递给后代(后代)的多细胞生物细胞群。 换句话说,它们是形成卵子、精子和受精卵的细胞。 它们通常被分化以执行此功能,并被隔离在远离其他身体细胞的特定位置。
通常,这种传递是通过有性生殖过程发生的;通常,它是一个过程,包括遗传物质的系统变化,例如在重组、减数分裂和受精过程中发生的变化。然而,多细胞生物有许多例外,包括各种形式的无性生殖、自体生殖、自体生殖、克隆或孤雌生殖等过程和概念。
种系细胞称为生殖细胞。例如,精子和卵子等配子是生殖细胞。分裂产生配子的细胞,称为配子体,产生配子的细胞,称为配子原细胞,一直回到受精卵,个体发育的细胞。
在有性繁殖的生物体中,不属于种系的细胞称为体细胞。根据这种观点,种系中的突变、重组和其他遗传变化可能会传递给后代,但体细胞中的变化不会。这不一定适用于体细胞繁殖的生物,例如一些多孔动物和许多植物。例如,当体细胞二倍体细胞取代胚珠或早期胚胎时,许多柑橘属植物、蔷薇科植物和一些菊科植物(如蒲公英)无融合生殖地产生种子。
在遗传思想的早期阶段,生殖细胞和体细胞之间有明显的区别。例如,奥古斯特·魏斯曼 (August Weismann) 提出并指出,生殖系细胞是不朽的,因为它是自生命开始以来无限繁殖的谱系的一部分,并且除非发生意外,否则可以无限期地继续繁殖。
然而,现在已详细了解体细胞和生殖细胞之间的这种区别部分是人为的,取决于特定情况和内部细胞机制,例如端粒和控制,例如端粒酶在生殖细胞、干细胞等中的选择性应用。
并非所有的多细胞生物都分化为体细胞和种系,但在缺乏专门的技术人为干预的情况下,除了最简单的多细胞结构外,几乎所有的多细胞生物都会分化。
在这样的生物体中,体细胞实际上往往是全能的,并且一个多世纪以来,人们都知道海绵细胞在通过筛子分离后会重新组装成新的海绵。
生命细细胞系可以指跨越多代个体的细胞谱系——例如,将任何活的个体与假设的最后一个普遍共同祖先联系起来的种系,所有植物和动物都起源于此。
进化
编辑植物和基础后生动物,如海绵 (Porifera) 和珊瑚 (Anthozoa) 不会隔离不同的种系,从多能干细胞谱系中产生配子,这些干细胞谱系也会产生普通的体细胞组织。因此,种系隔离很可能首先在具有复杂身体计划的复杂动物中进化,即两侧对称动物。
关于严格区分种系-体细胞的起源有几种理论。
在胚胎发生早期将孤立的生殖细胞群放在一边可能会促进复杂多细胞生物的体细胞之间的合作。
另一个最近的理论表明,早期种系隔离的进化是为了限制具有高能量需求和快速线粒体突变率的复杂生物体中线粒体基因有害突变的积累。
DNA损伤、突变与修复
编辑活性氧 (ROS) 作为新陈代谢的副产物产生。在种系细胞中,ROS 可能是 DNA 损伤的重要原因,DNA 损伤在 DNA 复制时会导致突变。
8-Oxoguanine 是鸟嘌呤的一种氧化衍生物,由小鼠生殖系细胞的自发氧化产生,在细胞的 DNA 复制过程中会导致 GC 到 TA 的颠换突变。
这种突变发生在整个小鼠染色体以及配子发生的不同阶段。
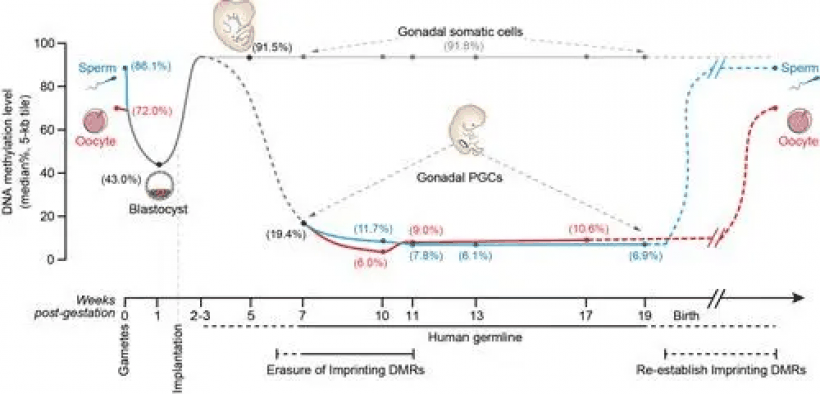

对于精子发生和卵子发生,配子发生不同阶段的细胞突变频率比体细胞低约 5 到 10 倍。与体细胞相比,种系细胞的突变频率较低似乎是由于在种系减数分裂过程中更有效地修复 DNA 损伤,特别是同源重组修复。
在人类中,大约 5% 的活产后代患有遗传疾病,其中约 20% 是由新出现的种系突变引起的。
表观遗传改变
编辑DNA 的表观遗传改变包括影响基因表达的修饰。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228931/