
CDNA文库
编辑CDNA 文库是插入宿主细胞集合中的克隆 cDNA(互补 DNA)片段的组合,构成生物体转录组的一部分,并作为文库存储。 cDNA 是由细胞核中发现的完全转录的 MRNA 产生的,因此仅包含生物体的表达基因。 同样,可以生成组织特异性 cDNA 文库。 在真核细胞中,成熟的 mRNA 已经被剪接,因此产生的 cDNA 没有内含子,可以很容易地在细菌细胞中表达。 虽然 cDNA 文库中的信息是一种强大而有用的工具,因为基因产物很容易识别,但文库缺乏有关增强子、内含子和基因组 DNA 文库中发现的其他调控元件的信息。
cDNA文库构建
编辑cDNA 是使用逆转录酶从真核细胞的成熟 mRNA 创建的。 在真核生物中,poly-(A) 尾巴(由长序列的腺嘌呤核苷酸组成)将 mRNA 与 tRNA 和 rRNA 区分开来,因此可用作逆转录的引物位点。 这有一个问题,即并非所有的转录本,例如组蛋白的转录本,都编码 poly-A 尾巴。
mRNA提取
编辑首先,需要分离 mRNA 模板以创建 cDNA 文库。 由于 mRNA 仅包含外显子,因此应考虑分离的 mRNA 的完整性,以便仍然可以生产编码的蛋白质。 分离的 mRNA 应在 500 bp 到 8 kb 之间。 存在几种纯化 RNA 的方法,例如三唑提取和柱纯化。 可以使用寡聚 dT 核苷酸包被树脂进行柱纯化,并且可以利用 mRNA 的特征,例如具有 poly-A 尾巴,其中只有包含该特征的 mRNA 序列才会结合。 然后洗脱与柱结合的所需 mRNA。
cDNA构建
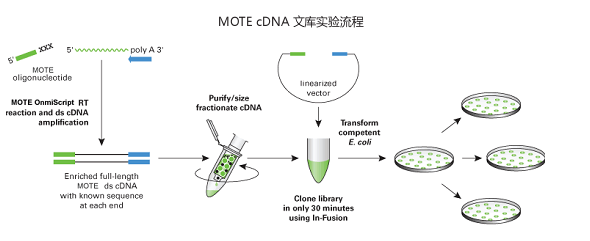
编辑一旦 mRNA 被纯化,oligo-dT 引物(脱氧胸苷核苷酸的短序列)就会与 RNA 的 poly-A 尾巴结合。 需要引物来启动逆转录酶的 DNA 合成。 这导致了 RNA-DNA 杂交体的产生,其中互补 DNA 的单链与 mRNA 链结合。 为了去除 mRNA,使用 RNAse H 酶切割 mRNA 的骨架并产生游离的 3′-OH 基团,这对于用 DNA 替换 mRNA 很重要。 然后加入 DNA 聚合酶 I,裂解的 RNA 作为引物,DNA 聚合酶 I 可以识别并启动 RNA 核苷酸与 DNA 核苷酸的替换。 这是由 sscDNA 本身通过在 3′ 端盘绕自身提供的,产生发夹环。 聚合酶延伸3′-OH端,随后3’端环在S1核酸酶的剪切作用下打开。 然后使用限制性核酸内切酶和 DNA 连接酶将序列克隆到细菌质粒中。
然后选择克隆的细菌,通常是通过使用抗生素选择。 一旦被选中,就会创建细菌种群,随后可以对其进行培养和测序以编译 CDNA 文库。
cDNA 库使用
编辑在复制真核基因组时通常使用 cDNA 文库,因为信息量会减少以从文库中移除大量非编码区域。 cDNA 文库用于在原核生物中表达真核基因。 原核生物的 DNA 中没有内含子,因此不具备任何可以在转录过程中将其切除的酶。 cDNA没有内含子,因此可以在原核细胞中表达。 cDNA 文库在反向遗传学中最有用,在这种情况下,额外的基因组信息用处不大。 此外,cDNA 文库经常用于功能性克隆,以根据编码蛋白质的功能识别基因。 在研究真核 DNA 时,表达文库是使用互补 DNA (cDNA) 构建的,以帮助确保插入片段是真正的基因。
cDNA 文库与基因组 DNA 文库
编辑CDNA 文库缺乏基因组 DNA 中的非编码和调控元件。 基因组 DNA 文库提供了有关生物体的更多详细信息,但生成和保存需要更多资源。
cDNA克隆
编辑可以使用限制性位点接头克隆 cDNA 分子。 接头是短的双链 DNA(寡脱氧核糖核苷酸)片段,长约 8 至 12 个核苷酸对,包括限制性核酸内切酶切割位点。 cDNA 和接头都有平端,可以使用高浓度的 T4 DNA 连接酶将其连接在一起。 然后通过用适当的核酸内切酶切割 cDNA 末端(现在具有带有掺入位点的接头),在 cDNA 分子中产生粘性末端。 然后克隆载体(质粒)也被适当的核酸内切酶切割。 在将插入物的粘性末端连接到载体中之后,将所得的重组 DNA 分子转移到大肠杆菌宿主细胞中进行克隆。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229244/