
简介
编辑异染色质是一种紧密堆积的 DNA 或浓缩 DNA,有多种形式。 这些变体介于组成型异染色质和兼性异染色质这两个极端之间。 两者都在基因表达中发挥作用。 因为它是紧密堆积的,所以被认为聚合酶无法接近,因此不会被转录; 然而,根据 Volpe 等人的说法。 (2002),以及此后的许多其他论文,实际上大部分 DNA 都被转录了,但它通过 RNA 诱导的转录沉默 (RITS) 不断翻转。 最近对电子显微镜和 OsO4 染色的研究表明,致密堆积不是由染色质引起的。
组成型异染色质可以影响自身附近的基因(例如位置效应杂色)。 它通常是重复的并形成结构功能,例如着丝粒或端粒,此外还充当其他基因表达或抑制信号的吸引子。
兼性异染色质是基因通过组蛋白去乙酰化或通过 RNAi 的 Piwi 相互作用 RNA (piRNA) 等机制沉默的结果。 它不重复,具有组成型异染色质的紧凑结构。 然而,在特定的发育或环境信号提示下,它可能会失去其浓缩结构并变得具有转录活性。
异染颜色与人类基因组某些部分中 H3K9 的二甲基化和三甲基化有关。 H3K9me3 相关的甲基转移酶似乎在器官发生开始时的谱系定型过程中修饰异染色质和维持谱系保真度方面起着关键作用。
结构
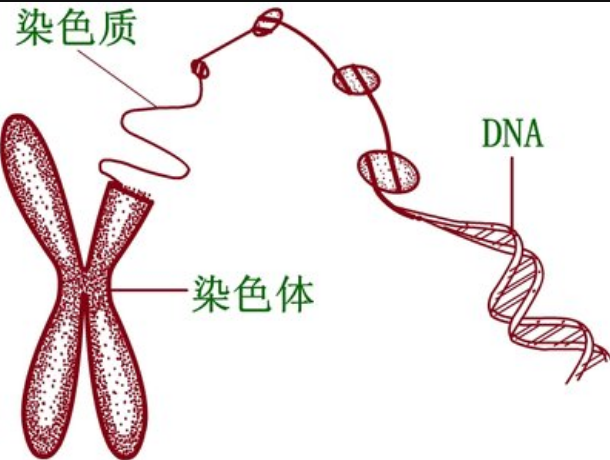
编辑染色质有两种类型:常染色质和异染色质。 最初,这两种形式在细胞学上的区别在于它们被染色的强度——常染色质强度较低,而异染色质染色强度较高,表明包装更紧密。 异染颜色通常局限于细胞核的外围。尽管有这种早期的二分法,但最近在动物和植物中的证据表明存在两种以上不同的异染色质状态,实际上可能存在四种或五种状态 ,每个都由不同的表观遗传标记组合标记。
异染色质主要由遗传失活的卫星序列组成,许多基因受到不同程度的抑制,尽管有些基因根本无法在常染色质中表达。 着丝粒和端粒都是异色的,女性中第二条失活的 X 染色体的 Barr 体也是异色的。
函数
编辑异染色素与多种功能有关,从基因调控到保护染色体完整性; 其中一些作用可归因于 DNA 的密集堆积,这使得通常结合 DNA 或其相关因子的蛋白质因子不易接近。 例如,裸露的双链 DNA 末端通常会被细胞解释为受损或病毒 DNA,从而触发细胞周期停滞、DNA 修复或片段破坏,例如通过细菌中的核酸内切酶。
染色质的某些区域非常密集地排列着纤维,这些纤维显示出与有丝分裂时染色体的状态相当的状态。 异染颜色一般是克隆遗传的; 当一个细胞分裂时,两个子细胞通常在相同的 DNA 区域内含有异染色质,从而导致表观遗传。 变异导致异染色质侵占相邻基因或从域极端的基因中退去。 可转录材料可以通过定位(顺式)在这些边界域来抑制。 这导致表达水平因细胞而异,这可以通过位置效应杂色来证明。 在组成型异染色质和高活性基因并列的罕见情况下,绝缘子序列可能充当屏障(例如,鸡 β-珠蛋白基因座上游的 5’HS4 绝缘子,以及两个酵母属中的基因座)。
组成型异染色质
编辑给定物种的所有细胞都在组成型异染色质中包装相同的 DNA 区域,因此在所有细胞中,组成型异染色质中包含的任何基因都将表达不良。 例如,所有人类染色体 1、9、16 和 Y 染色体都包含大的组成型异染色质区域。 在大多数生物体中,组成型异染色质发生在染色体着丝粒周围和端粒附近。
兼性异染色质
编辑包装在兼性异染色质中的 DNA 区域在一个物种内的细胞类型之间将不一致,因此一个细胞中包装在兼性异染色质中的序列(并且其中的基因表达很差)可能会包装在另一个细胞的常染色质中 (并且其中的基因不再沉默)。 然而,兼性异染色质的形成受到调节,并且通常与形态发生或分化有关。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/250152/