
真核起始因子
编辑真核起始因子 (eIF) 是参与真核翻译起始阶段的蛋白质或蛋白质复合物。 这些蛋白质有助于稳定起始密码子周围核糖体预起始复合物的形成,并且是转录后基因调控的重要输入。 几个起始因子与小的 40S 核糖体亚基和 Met-tRNAiMet 形成复合物,称为 43S 预起始复合物 (43S PIC)。 eIF4F 复合物的其他因子(eIF4A、E 和 G)将 43S PIC 募集到 MRNA 的五质帽结构,43S 粒子从该结构沿着 mRNA 扫描 5′–>3′ 到达 AUG 起始密码子。 Met-tRNAiMet 对起始密码子的识别促进了门控磷酸盐和 eIF1 的释放以形成 48S 预起始复合物 (48S PIC),随后募集 60S 核糖体大亚基以形成 80S 核糖体。 真核起始因子比原核起始因子多得多,反映出真核翻译的生物学复杂性更高。 至少有 12 种真核起始因子,由更多的多肽组成。
eIF1 和 eIF1A
编辑eIF1 和 eIF1A 都与 40S 核糖体亚基-mRNA 复合物结合。 它们一起诱导 mRNA 结合通道的开放构象,这对于扫描、tRNA 传递和启动密码子识别至关重要。 特别是,eIF1 从 40S 亚基解离被认为是起始密码子识别的关键步骤。eIF1 和 eIF1A 是小蛋白质(在人类中分别为 13 和 16 kDa)并且都是 43S PIC 的组成部分。 eIF1 在核糖体 P 位点附近结合,而 eIF1A 在 A 位点附近结合,其方式分别类似于结构和功能相关的细菌对应物 IF3 和 IF1。
eIF2
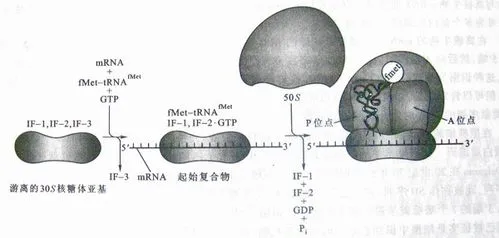
编辑eIF2 是主要的蛋白质复合物,负责将启动子 tRNA 递送到预启动复合物的 P 位点,作为包含 Met-tRNAiMet 和 GTP 的三元复合物(eIF2-TC)。 eIF2 对带蛋氨酸的起始 tRNA 具有特异性,这与用于延长多肽链的带蛋氨酸的其他 tRNA 不同。 eIF2 三元复合物仍然与 P 位点结合,而 mRNA 附着在 40s 核糖体上,复合物开始扫描 mRNA。 一旦 AUG 起始密码子被识别并位于 P 位点,eIF5 就会刺激 eIF2-GTP 的水解,通过门控磷酸盐释放有效地将其转换为 GDP 结合形式。 eIF2-GTP 的水解提供了构象变化,将扫描复合物变为 48S 起始复合物,引发剂 tRNA-Met 反密码子碱基与 AUG 配对。 起始复合物形成后,60s 亚基加入,eIF2 连同大多数起始因子从复合物中解离,允许 60S 亚基结合。 eIF1A 和 eIF5-GTP 在 A 位点保持相互结合,必须水解才能释放并正确启动伸长。
eIF2 具有三个亚基,即 eIF2-α、β 和 γ。 前 α-亚基是调节磷酸化的靶标,对于可能需要全局关闭蛋白质合成作为对细胞信号事件的反应的细胞特别重要。 当磷酸化时,它隔离 eIF2B(不要与 eIF2β 混淆),一种 GEF。 没有这个 GEF,GDP 就不能换成 GTP,翻译也就被打压了。 这方面的一个例子是 eIF2α 诱导的翻译抑制,这种抑制发生在网织红细胞缺铁时。 在病毒感染的情况下,当在许多多细胞生物中检测到 dsRNA 时,蛋白激酶 R (PKR) 会磷酸化 eIF2α,从而导致细胞死亡。
蛋白质 eIF2A 和 eIF2D 在技术上都被命名为“eIF2”,但它们都不是 eIF2 异源三聚体的一部分,它们似乎在翻译中发挥独特的功能。 相反,它们似乎分别参与了专门的途径,例如“eIF2 独立”翻译启动或重新启动。
eIF3
编辑eIF3 独立结合 40S 核糖体亚基、多个起始因子以及细胞和病毒 mRNA。
在哺乳动物中,eIF3 是最大的起始因子,由 13 个亚基 (a-m) 组成。 它的分子量约为 800 kDa,并控制 40S 核糖体亚基在具有 5′ 帽或 IRES 的 mRNA 上的组装。
eIF3 可以使用 eIF4F 复合物,或者在内部启动期间使用 IRES,将 mRNA 链定位在 40S 核糖体亚基的出口位点附近,从而促进功能性预启动复合物的组装。
在许多人类癌症中,eIF3 亚基过度表达(亚基 a、b、c、h、i 和 m)和表达不足(亚基 e 和 f)。 解释这种失调的一个潜在机制来自以下发现:eIF3 结合一组特定的细胞增殖调节因子 mRNA 转录物并调节它们的翻译。 eIF3 还通过 S6K1 和 mTOR/Raptor 介导细胞信号传导以影响翻译调节。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229291/