
真菌繁殖
编辑真菌繁殖是一个由交配类型决定的复杂过程。 真菌交配的研究主要集中在几种具有不同行为的模式物种上。 并非所有的真菌都进行有性繁殖,而且许多真菌都是等配的; 因此,对于真菌王国的许多成员来说,男性和女性这两个词并不适用。 同宗物种能够与自己交配,而在异宗物种中,只有相反交配类型的分离株才能交配。
同配真菌之间的交配可能仅包括将细胞核从一个细胞转移到另一个细胞。 物种内的营养不相容性通常会阻止一种真菌分离物与另一种分离物交配。 同一不相容群的分离株不会交配或交配不会产生成功的后代。 据报道,高度变异包括相同的化学型交配、孢子体到配子体的交配和线粒体的双亲转移。
在接合菌中交配
编辑接合菌菌丝向相容的配偶生长,它们都形成一座桥,称为原配子体,通过胞浆配子在菌丝尖端连接。 一对隔膜围绕合并的尖端形成,包围来自两个分离物的细胞核。 第二对隔膜形成两个相邻的单元格,每侧一个。 这些称为悬索的相邻细胞提供结构支撑。 中央细胞注定要成为孢子。 细胞核加入称为核配子的过程以形成受精卵。
在子囊菌中交配
编辑当它接近配偶时,单倍体囊真菌会发育出两个互补器官之一,即雌性子囊菌或雄性花囊。 这些器官类似于配子体,只是它们只包含细胞核。 一座桥,毛状体形成,为细胞核提供了从花药壳到子囊壳的通道。 双核生物从子囊长出,核配子发生在子实体中。
粗糙链孢霉
编辑粗糙脉孢菌是子囊菌门的一种红面包霉菌。 N. crassa 被用作模式生物,因为它易于生长并且具有单倍体生命周期:这使得遗传分析变得简单,因为隐性特征会在后代中出现。 减数分裂产物在称为子囊 (pl. asci) 的囊状结构中的有序排列有助于遗传重组的分析。 在自然环境中,粗糙猪笼草主要生活在热带和亚热带地区。 它经常可以在火灾后的死植物物质上生长。
Edward Tatum 和 George Wells Beadle 在他们于 1958 年获得诺贝尔生理学或医学奖的实验中使用了脉孢菌。这些实验的结果直接导致了一种基因,一种酶假说,即特定基因编码特定蛋白质。 这个概念启动了分子生物学。 只有当两个不同交配类型的细胞聚集在一起时,才能形成有性子实体(perithecia)。 与其他子囊菌一样,N. crassa 有两种交配类型,在这种情况下,用 A 和 a 表示。 A和a交配型菌株之间没有明显的形态差异。 两者都能形成丰富的protoperithecia,即雌性生殖结构。 当在氮源相对较低的固体(琼脂)合成培养基上生长时,原包皮细胞在实验室中最容易形成。 氮饥饿似乎是参与性发育的基因表达所必需的。 protoperithecium 由 ascogonium 组成,这是一种盘绕的多细胞菌丝,包裹在菌丝的结状聚集体中。
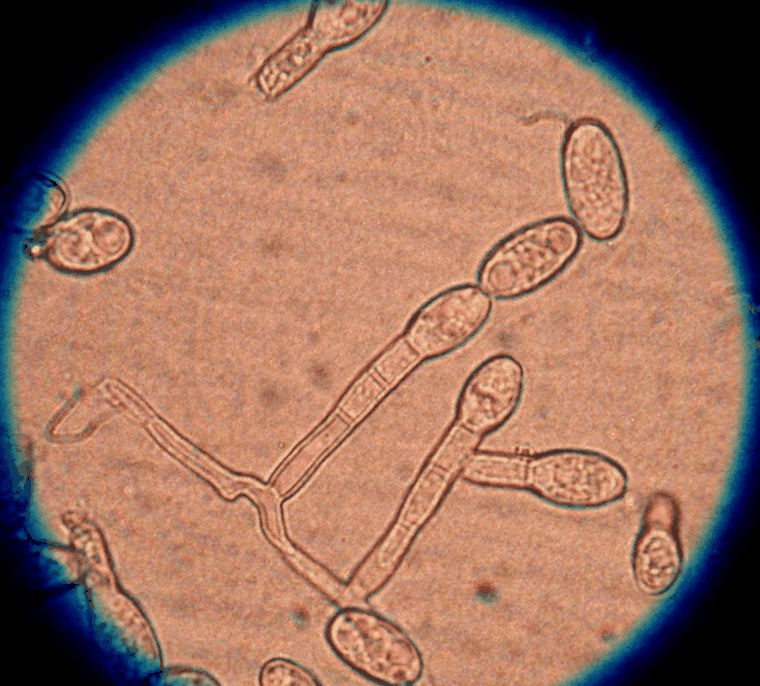
细长菌丝的分支系统,称为毛状体,从子囊的尖端伸出,超出覆盖的菌丝进入空气中。 当相反交配类型的细胞(通常是分生孢子)接触到毛的一部分时,性周期开始(即发生受精)。 这种接触之后可以进行细胞融合,导致来自受精细胞的一个或多个细胞核向下迁移到子囊细胞中。 由于 A 和 a 品系具有相同的性结构,因此不能将这两种品系视为完全雄性或雌性。 然而,作为接受者,A 和 a 菌株的原包皮都可以被认为是雌性结构,而受精分生孢子可以被认为是雄性参与者。
FINCHam 和 Day 概述了 A 和单倍体细胞融合后的后续步骤。 瓦格纳和米切尔。 细胞融合后,细胞核的进一步融合被延迟。 相反,受精细胞的细胞核和子囊细胞的细胞核结合在一起并开始同步分裂。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229392/