内源性反转录病毒
编辑内源性反转录病毒 (ERV) 是基因组中的内源性病毒元件,与逆转录病毒非常相似并可衍生自逆转录病毒。 它们在有颌脊椎动物的基因组中含量丰富,占人类基因组的 5-8%(较低估计约为 1%)。
ERV 是一种垂直遗传的前病毒序列,是一种称为转座子的基因的子类,转座子通常可以在基因组中包装和移动,在基因表达和调控中发挥重要作用。 然而,ERV 缺乏大多数转座子功能,通常不具有传染性,并且通常是逆转录病毒复制周期的有缺陷的基因组残余物。 由于它们整合并逆转录到宿主细胞的核基因组中,它们被区分为种系原病毒逆转录元件。
研究人员认为,逆转录病毒是从一种称为逆转录转座子的转座子进化而来的,它是一种 I 类元件; 这些基因可以发生突变,而不是移动到基因组中的另一个位置,它们可以变成外源性或致病性的。 这意味着并非所有 ERV 都起源于逆转录病毒的插入,但有些可能是它们相似的逆转录病毒中遗传信息的来源。 当病毒 DNA 的整合发生在种系中时,它会产生 ERV,后者随后会固定在宿主种群的基因库中。
编队
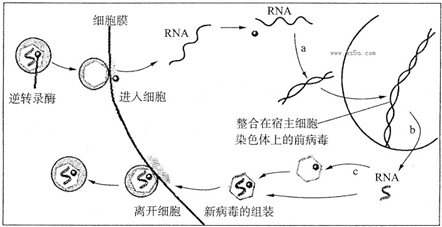
编辑逆转录病毒的复制周期需要将病毒基因组的 DNA 拷贝插入(整合)到宿主细胞的核基因组中。 大多数逆转录病毒感染体细胞,但偶尔也会感染生殖细胞(产生卵子和精子的细胞)。 极少数情况下,逆转录病毒整合可能发生在继续发育成活生物体的种系细胞中。 这种生物体将携带插入的逆转录病毒基因组作为其自身基因组的组成部分——一种内源性逆转录病毒 (ERV),可能作为新的等位基因由其后代遗传。 许多 ERV 已在其宿主的基因组中存在数百万年。 然而,其中大部分在宿主 DNA 复制过程中发生了失活突变,不再能够产生病毒。 ERV 也可以通过称为重组删除的过程从基因组中部分切除,其中新整合的逆转录病毒侧翼的相同序列之间的重组导致病毒基因组内部蛋白质编码区域的删除。
一般的逆转录病毒基因组由三个对其病毒基因组的入侵、复制、逃逸和传播至关重要的基因组成。 这三个基因是 gag(编码病毒核心的结构蛋白)、pol(编码逆转录酶、整合酶和蛋白酶)和 env(编码病毒外部的外壳蛋白)。 这些病毒蛋白被编码为多蛋白。 为了执行它们的生命周期,逆转录病毒严重依赖宿主细胞的机制。 蛋白酶降解病毒多蛋白的肽键,使分离的蛋白质发挥作用。 逆转录酶的作用是在病毒进入细胞核之前从宿主细胞细胞质中的病毒 RNA 合成病毒 DNA。 整合酶指导病毒 DNA 整合到宿主基因组中。
随着时间的推移,ERVs的基因组不仅会发生点突变,还会与其他ERVs发生洗牌和重组。 环境序列衰减的 ERV 变得更有可能传播。
在基因组进化中的作用
编辑内源性反转录病毒可以在塑造基因组方面发挥积极作用。 该领域的大多数研究都集中在人类和高等灵长类动物的基因组上,但也对其他脊椎动物,如小鼠和绵羊进行了深入研究。 位于 ERV 基因组侧翼的长末端重复序列 (LTR) 序列经常充当替代启动子和增强子,通常通过产生组织特异性变异来促进转录组。 此外,逆转录病毒蛋白本身已被增选用于新的宿主功能,特别是在繁殖和发育方面。 同源逆转录病毒序列之间的重组也有助于基因改组和遗传变异的产生。 此外,在逆转录病毒序列的潜在拮抗作用的情况下,阻遏基因共同进化以对抗它们。

大约 90% 的内源性逆转录病毒是单独的 LTR,缺少所有开放阅读框 (ORF)。 Solo LTR 和与完整逆转录病毒序列相关的 LTR 已被证明可作为宿主基因的转录元件。 它们的作用范围主要是插入到蛋白质编码基因的5'UTR; 然而,众所周知,它们会作用于距离最远 70-100 kb 的基因。 这些元件中的大多数以有义方向插入到它们相应的基因中,但有证据表明 LTR 以反义方向起作用并作为相邻基因的双向启动子。 在少数情况下,LTR 充当该基因的主要启动子。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229309/