
内质网
编辑内质网(ER)是一种类型的细胞器两个亚基组成- 粗面内质网(RER)和滑面内质网(SER)。内质网存在于大多数真核细胞中,并形成扁平的,膜包裹的囊(在RER中被称为池)和SER中的管状结构的互连网络。ER的膜与核外膜是连续的。内质网未见于红细胞或精子。
两种类型的ER共享许多相同的蛋白质,并从事某些共同的活动,例如某些脂质和胆固醇的合成。取决于细胞的活性,不同类型的细胞包含两种类型的ER的不同比率。
粗糙的内质网的外表面(胞质)散布着核糖体,核糖体是蛋白质合成的部位。粗糙的内质网在诸如肝细胞的细胞中尤为突出。光滑的内质网缺乏核糖体,在脂质合成中起作用,但没有代谢,没有甾体激素的产生和排毒。平滑的内质网在哺乳动物的肝脏和性腺细胞中尤为丰富。
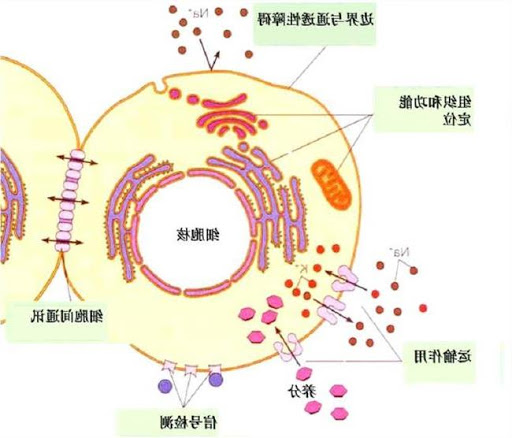
内质网的结构
编辑内质网的总体结构是称为池的膜网络。这些囊状结构通过细胞骨架结合在一起。的磷脂膜包围脑池空间,其是与连续核周空间而是从单独的胞质溶胶。内质网的功能可以概括为蛋白质和膜脂质的合成和输出,但在ER与细胞类型和细胞功能之间有所不同。细胞中粗糙和光滑的内质网的数量可以缓慢地从一种转变为另一种,这取决于细胞的代谢活性。转化可包括在膜中嵌入新蛋白以及结构变化。蛋白质含量可能发生变化,而没有明显的结构变化。
粗面内质网
编辑粗糙的内质网(通常缩写为RER或粗糙的ER;也称为颗粒状内质网)的表面布满了蛋白质制造的核糖体,使其外观呈“粗糙”(因此得名)。核糖体在粗糙内质网上的结合位点是translocon。然而,核糖体并不是细胞器结构的稳定部分,因为它们不断被结合并从膜中释放出来。核糖体仅在细胞质中形成特定的蛋白质-核酸复合物后才与RER结合。当游离核糖体开始这种特殊的复杂形式翻译的一种分泌途径的蛋白质的MRNA。聚合的前5-30 个氨基酸编码信号肽,一种被信号识别颗粒(SRP)识别并结合的分子信息。翻译暂停和核糖体的复合物结合到RER 易位子其中翻译继续与新生的(新)蛋白质形成到RER内腔和/或膜。该蛋白质在内腔中被一种酶(一种信号肽酶),从而去除信号肽。此时,核糖体可能会释放回细胞质中。然而,非翻译的核糖体也与转座子保持联系。
粗糙的内质网的膜形成大的双膜片,其位于核包膜的外层附近并与其连续。双层膜片堆叠并通过几个右旋或左旋螺旋坡道(“ Terasaki坡道”)连接,形成了一个类似于多层停车场的结构。虽然在内质网和高尔基体之间没有连续的膜,但膜结合的运输小泡在这两个区室之间穿梭蛋白质。囊泡被称为COPI和COPII的包膜蛋白包围。COPII将囊泡靶向高尔基体,而COPI则将其带回到粗糙的内质网。在演唱会的粗面内质网的工作原理与高尔基复合体,以针对新的蛋白质,以正确的目的地。转运出内质网的第二种方法涉及称为膜接触位点的区域,内质网和其他细胞器的膜紧密结合在一起,从而可以转运脂质和其他小分子。
粗糙的内质网是多种功能的关键:
- 在顺式-高尔基体网络中添加带有甘露糖6-磷酸酯标记的溶酶体酶。
- 分泌蛋白的生产,要么无标签地组成性分泌,要么以调控方式分泌,其中涉及网格蛋白和信号肽中的配对碱性氨基酸。
- 整合膜蛋白是保持嵌在膜的囊泡退出并绑定到新的膜。Rab蛋白是靶向膜的关键。SNAP和SNARE蛋白是融合事件的关键。
- 随着组装的继续,初始糖基化。这是N链接的(高尔基体中发生O链接)。
平滑内质网
编辑在大多数细胞中,缺乏光滑的内质网(缩写为SER)。取而代之的是,ER的一部分区域平滑且部分粗糙,该区域称为过渡ER。过渡ER之所以得名,是因为它包含ER出口站点。在这些区域中,含有在ER中产生的脂质和蛋白质的运输囊泡与ER分离,并开始向高尔基体移动。特化细胞可具有许多平滑的内质网,并且在这些细胞中,平滑的ER具有许多功能。它综合脂质、磷脂和类固醇。分泌这些产物的细胞,例如睾丸、卵巢和皮脂腺中的细胞,具有丰富的平滑内质网。它还进行碳水化合物的代谢,天然代谢产物以及酒精和药物的解毒,细胞膜蛋白上的受体附着以及类固醇代谢。在肌肉细胞中,它调节钙离子浓度。平滑内质网存在于多种细胞类型(动物和植物)中,并且在每种细胞中均具有不同的功能。光滑的内质网还包含葡萄糖6磷酸酶,将6-磷酸葡萄糖转化为葡萄糖,这是糖异生的一个步骤。它连接到核被膜,由位于细胞外围附近的小管组成。这些管有时分支形成外观呈网状的网络。在某些细胞中,有扩张的区域,如粗糙的内质网囊。光滑的内质网网络允许增加表面积用于关键酶和这些酶产物的作用或储存。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/114030/