
λ噘菌体
编辑(了解如何以及何时删除此模板消息)
肠杆菌噬菌体 λ(λ 噬菌体、大肠杆菌噬菌体 λ,正式名称为大肠杆菌λ 病毒)是一种细菌病毒或噬菌体,可感染大肠杆菌(大肠杆菌)。 它是由 Esther Lederberg 于 1950 年发现的。这种病毒的野生型具有温和的生命周期,这使得它可以通过溶源性驻留在宿主的基因组中或进入裂解期,在此期间它会杀死并裂解细胞以 产生后代。 在特定位点发生突变的 Lambda 菌株无法溶原细胞; 相反,它们在重复感染已经溶原化的细胞后生长并进入裂解周期。
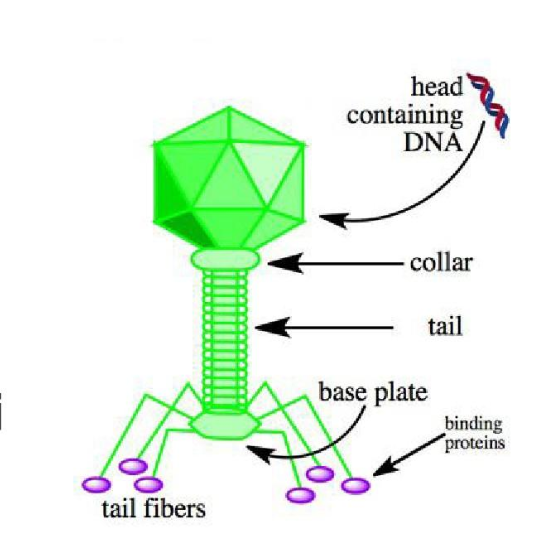
噬菌体颗粒由头部(也称为衣壳)、尾巴和尾纤维组成(见下图病毒)。 头部包含噬菌体的双链线性 DNA 基因组。 在感染过程中,噬菌体颗粒识别并与其宿主大肠杆菌结合,导致噬菌体头部的 DNA 通过尾部喷射到细菌细胞的细胞质中。 通常,随后会发生裂解循环,其中 lambda DNA 被复制并在细胞内产生新的噬菌体颗粒。 随后是细胞裂解,将细胞内容物(包括已组装的病毒体)释放到环境中。 然而,在某些条件下,噬菌体 DNA 可能会通过溶原途径将自身整合到宿主细胞染色体中。 在这种状态下,λ DNA 被称为原噬菌体,并驻留在宿主的基因组中,对宿主没有明显的伤害。 当存在原噬菌体时,宿主被称为溶原体。 当溶源菌进入应激条件时,这种前噬菌体可能会进入裂解周期。
解剖学
编辑病毒颗粒由头部和尾部组成,尾部可以有尾纤维。 整个颗粒由 12-14 种不同的蛋白质组成,总共有超过 1000 个蛋白质分子和一个位于噬菌体头部的 DNA 分子。 然而,目前还不完全清楚 L 和 M 蛋白是否是病毒体的一部分。 所有表征的 lambdoid 噬菌体都具有 N 蛋白介导的转录抗终止机制,但噬菌体 HK022 除外
基因组包含 48,502 个碱基对的双链线性 DNA,在两个 5′ 末端都有 12 个碱基的单链片段。 这两个单链片段是所谓的 cos 位点的粘端。 cos 位点使宿主细胞质中的 DNA 环化。 因此,在其环状形式中,噬菌体基因组的长度为 48,502 个碱基对。 lambda 基因组可以插入大肠杆菌染色体,然后称为原噬菌体。 有关详细信息,请参阅下面的部分。
生命周期
编辑感染
编辑λ 噬菌体是一种非收缩性尾噬菌体,这意味着在感染事件中它不能“迫使”其 DNA 穿过细菌细胞膜。 它必须改用现有的途径来侵入宿主细胞,进化出尾巴的尖端以与特定的孔相互作用,以允许其 DNA 进入宿主。
- 噬菌体 Lambda 通过尾端的 J 蛋白与大肠杆菌细胞结合。 J蛋白与大肠杆菌的麦芽糖外膜孔蛋白(lamB基因的产物)相互作用,该孔蛋白分子是麦芽糖操纵子的一部分。
- 线性噬菌体基因组通过外膜注入。
- DNA 穿过内膜中的甘露糖通透酶复合物(由 manXYZ 基因编码),并立即使用 cos 位点、富含 12 个碱基的 G-C 粘性末端进行环化。 单链病毒 DNA 末端由宿主 DNA 连接酶连接。 人们普遍不认为 12 bp λ 粘性末端是生物 DNA 的第一个直接核苷酸测序的主题。
- 宿主 DNA 旋转酶将负超螺旋置于环状染色体中,导致富含 A-T 的区域解旋并驱动转录。
- 转录从产生“立即早期”转录本的组成型 PL、PR 和 PR 启动子开始。 首先,它们表达 N 和 cro 基因,产生 N、Cro 和一种短的无活性蛋白。
- Cro 与 OR3 结合,阻止进入 PRM 启动子,阻止 cI 基因的表达。 N与两个Nut(N利用)位点结合,一个在PL阅读框的N基因中,一个在PR阅读框的cro基因中。
- N 蛋白是一种抗终止剂,通过在新生转录的 MRNA 的特定位点结合转录 RNA 聚合酶发挥作用。 当 RNA 聚合酶转录这些区域时,它会募集 N 并与几种宿主 Nus 蛋白形成复合物。 这个复合体会跳过大多数终止序列。 扩展转录本(“晚早”转录本)包括 N 和 cro 基因以及 cII 和 cIII 基因,以及稍后讨论的 xis、int、O、P 和 Q 基因。
- cIII 蛋白的作用是保护 cII 蛋白免受 FtsH(一种膜结合的必需大肠杆菌蛋白酶)的蛋白水解。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228373/