
限制酶
编辑限制酶e,更准确地说是限制性核酸内切酶(REN),是可以在特定位置识别和切割DNA的酶。 限制性核酸内切酶存在于细菌和古细菌等中,用于抵御噬菌体。 限制酶 e 通过缺失的甲基化模式或通过其他不存在的 DNA 序列识别外来 DNA,然后水解外来 DNA。 因此,它们总是与典型的 DNA 甲基转移酶一起出现在细菌中,后者在细菌自身的 DNA 上留下特征模式。
属性
编辑为了使细菌具有限制酶作为防御系统,至少需要三个功能上可区分的蛋白质区域:限制域、甲基化域和序列识别域。 这三个结构域可以只存在于一种蛋白质上,也可以分布在多个蛋白质上。
每种限制性核酸内切酶识别特定的 DNA 碱基序列。 限制酶在限制性消化中的特异性取决于所用的缓冲液和辅助因子等。
限制发酵剂在生物化学中用于在指定点切割 DNA,例如 B. 在限制性分析(限制性酶切后进行琼脂糖凝胶电泳)或克隆中。 因此,这些酶也被称为分子剪刀。 连接酶用于共价重新连接切割末端(在粘性末端的情况下仅在杂交后)。 如果环境条件不对,也可以施加不明确的限制,这就是所谓的明星活动。 限制性核酸内切酶的特异性可以通过蛋白质设计特别适用于所需的 DNA 序列,例如 B. 在锌指核酸酶中。
个体限制酶的界面位置可以用 DNA 的限制性图谱表示。 例如,存在这样的基因组和质粒图谱。 当 DNA 被限制酶 e 切割时产生的 DNA 片段的长度可用于与限制性图谱相比较来识别 DNA 部分。
分类
编辑根据它们的性质,主要有四种类型,又分为几个亚型:
- I 型在远离识别序列的随机位置切割 DNA。 需要 ATP 并从 S-腺苷甲硫氨酸转移甲基。
- II 型在识别序列内或附近切割 DNA。 不需要 ATP,也没有甲基转移酶活性。
- III 型将 DNA 切割离识别序列约 20 到 25 个碱基对。 需要 ATP 并从 S-腺苷甲硫氨酸转移甲基。
- IV 型仅切割甲基化/羟甲基化/葡糖基-羟甲基化 DNA – 与受甲基化模式抑制的 I-III 型相反。
- V 型使用向导 RNA来靶向入侵生物体上的特定非回文序列。 如果提供合适的向导 RNA,它可以切割可变长度的 DNA。 由于其灵活性和易用性,V 型限制发酵剂是未来基因工程应用的有前途的候选者。
II 型有已知的亚型 IIS 型和 IIG 型,它们切入识别序列之外。
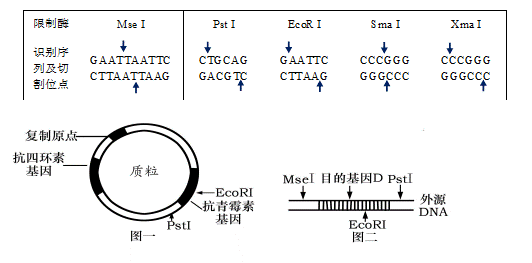
在一般用法中,术语“限制酶”通常等同于 II 型限制性核酸内切酶,因为 I 型和 III 型酶在分子生物学中的重要性不大。 限制发酵剂的名称表明了它们的来源。 第一个字母代表属,第二个和第三个字母代表种,它由名称后缀和发现的时间顺序扩展。 例如,EcoRI 酶是在大肠杆菌 R(粗)菌株中发现的第一种酶,而 SmaI 是粘质沙雷氏菌中的第一种酶。 具有相同识别序列和相同切割模式的不同来源的限制酶称为同裂酶。 如果它们在同一序列内切割但留下不同的切割末端,则它们被称为新裂酶。
II型限制性核酸内切酶的识别序列主要由4、6或8个碱基对的回文序列组成。 切口可以是直的。 这种平末端导致克隆过程中较低的连接产量。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/375959/