
β-氧化
编辑在生物化学和新陈代谢中,β-氧化是脂肪酸分子在原核生物的胞质溶胶和真核生物的线粒体中分解生成乙酰辅酶 A(进入柠檬酸循环)以及 NADH 和 FADH2(后者进入柠檬酸循环)的分解代谢过程。 是电子传递链中使用的辅酶。 之所以这样命名,是因为脂肪酸的 β 碳会氧化成羰基。 β-氧化主要由线粒体三功能蛋白促进,这是一种与线粒体内膜相关的酶复合物,尽管非常长的链脂肪酸在过氧化物酶体中被氧化。
激活和膜转运
编辑由于带负电荷,游离脂肪酸不能穿透任何生物膜。 游离脂肪酸必须通过特定的转运蛋白(例如 SLC27 家族脂肪酸转运蛋白)穿过细胞膜。 一旦进入胞质溶胶,以下过程会将脂肪酸带入线粒体基质,以便发生 β 氧化。
- 长链脂肪酸 – CoA 连接酶催化脂肪酸与 ATP 之间的反应,生成脂肪酰基腺苷酸和无机焦磷酸盐,无机焦磷酸盐随后与游离辅酶 A 反应生成脂肪酰基辅酶 A 酯和 AMP .
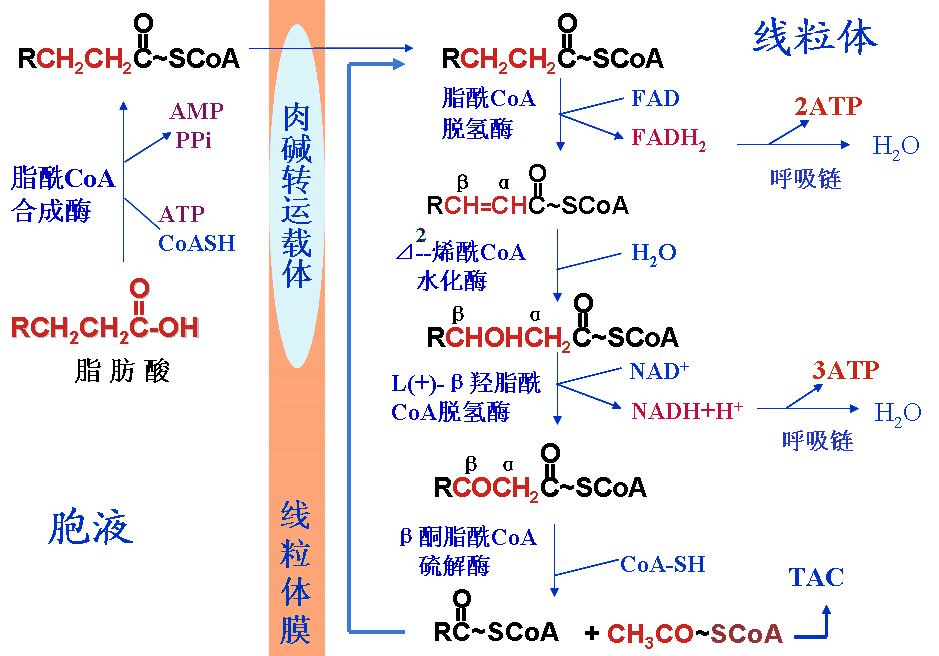
- 如果脂酰辅酶 A 具有长链,则必须利用肉碱穿梭:
- 酰基辅酶 A 通过肉碱棕榈酰转移酶 I 转移到肉碱的羟基,位于线粒体内外膜的胞质表面。
- 酰基肉碱通过肉碱-酰基肉碱转位酶在内部穿梭,而肉碱在外部穿梭。
- 酰基肉碱通过位于线粒体内膜内表面的肉碱棕榈酰转移酶 II 转化回酰基辅酶 A。 释放的肉碱穿梭回胞质溶胶,就像酰基肉碱穿梭到基质中一样。
- 如果脂肪酰基辅酶 A 含有短链,这些短链脂肪酸可以简单地扩散穿过线粒体内膜。
一般机制
编辑一旦脂肪酸进入线粒体基质内部,β-氧化就会通过在每个循环中裂解两个碳原子形成乙酰辅酶 A 来发生。 该过程包括 4 个步骤。
- 长链脂肪酸脱氢后在 C2 和 C3 之间形成反式双键。 这是由酰基辅酶 A 脱氢酶催化产生反式 δ 2-烯酰辅酶 A。 它以FAD为电子受体,被还原为FADH2。
- 反式 deltA2-烯酰辅酶 A 在双键处水合,通过烯酰辅酶 A 水合酶产生 L-3-羟酰基辅酶 A。
- L-3-羟酰基辅酶 A 再次脱氢,通过 3-羟酰基辅酶 A 脱氢酶生成 3-酮酰基辅酶 A。 该酶使用 NAD 作为电子受体。
- 硫解发生在 3-酮脂酰辅酶 A 的 C2 和 C3(α 和β 碳)之间。 当一个新的辅酶 A 分子通过对 C3 的亲核攻击断开键时,硫解酶会催化反应。 这释放了前两个碳单元,如乙酰辅酶 A 和脂肪酰基辅酶 A 减去两个碳。 该过程一直持续到脂肪酸中的所有碳都转化为乙酰辅酶 A。
脂肪酸被体内大部分组织氧化。 然而,一些组织,如哺乳动物的红细胞(不含线粒体)和中枢神经系统的细胞不使用脂肪酸来满足其能量需求,而是使用碳水化合物(红细胞和神经元)或酮 体(仅限神经元)。
由于许多脂肪酸未完全饱和或不具有偶数个碳,因此演化出几种不同的机制,如下所述。
偶数饱和脂肪酸
编辑一旦进入线粒体,每个 β-氧化循环都会释放一个双碳单元(乙酰辅酶 A),并依次发生四个反应:
这个过程一直持续到整个链被切割成乙酰辅酶 A 单元。 最后一个循环产生两个独立的乙酰辅酶 A,而不是一个酰基辅酶 A 和一个乙酰辅酶 A。 对于每个循环,酰基 CoA 单元都会缩短两个碳原子。 同时,形成一分子 FADH2、NADH 和乙酰辅酶 A。
奇数饱和脂肪酸
编辑通常,在植物和一些海洋生物的脂质中发现了碳原子数为奇数的脂肪酸。 许多反刍动物在发酵过程中会形成大量的3-碳丙酸。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228812/