
真核起始因子3
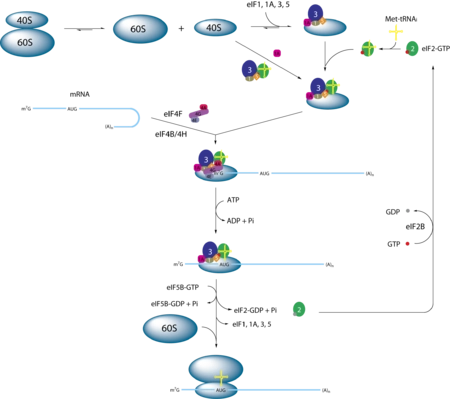
编辑真核起始原因3 (eIF3) 是一种多蛋白复合物,在真核翻译的起始阶段发挥作用。 它对于大多数形式的帽依赖和帽独立翻译启动是必不可少的。 在人类中,eIF3 由 13 个不同的亚基 (eIF3a-m) 组成,总分子量约为 800 kDa,使其成为最大的翻译起始因子。 eIF3 复合体在真核生物中广泛保守,但单个亚基的保守性因生物体而异。
函数
编辑eIF3 刺激翻译启动的几乎所有步骤。 eIF3 似乎也参与翻译的其他阶段,例如回收,它促进终止后核糖体的分裂。 在 uORF 后重新启动的特殊情况下,eIF3 可能通过延伸和终止与核糖体保持结合,以促进后续的启动事件。 研究还表明,eIF3 通过与终止前复合物相互作用并干扰解码,在酵母中的程序性终止密码子通读中发挥作用。
互动
编辑eIF3 在其溶剂侧及其附近结合小核糖体亚基 (40S),并充当其他几种起始因子、辅助因子 DHX29 和 MRNA 的支架。 eIF3 是多因子复合物 (MFC) 以及 43S 和 48S 预启动复合物 (PIC) 的组成部分。 eIF3 与其他启动因子的相互作用因物种而异; 例如,哺乳动物 eIF3 直接与 eIF4F 复合体相互作用(通过 eIF4G),而出芽酵母则缺乏这种联系。 然而,哺乳动物和酵母 eIF3 都独立地结合 eIF1、eIF4B 和 eIF5。
eIF3 的几个亚基包含 RNA 识别基序 (RRM) 和其他 RNA 结合结构域,形成多亚基 RNA 结合界面,eIF3 通过该界面与细胞和病毒 IRES mRNA(包括 HCV IRES)相互作用。 eIF3 还被证明可以特异性结合 5’UTR 内的 m6A 修饰的 RNA,以促进不依赖于帽的翻译。
出芽酵母 eIF3 的所有五个核心亚基以及其他几个翻译因子都存在于热诱导应激颗粒中。
结构
编辑功能性 eIF3 复合物可以从天然资源中纯化,或从重组表达的亚基中重建。 单个亚基的结构已通过 X 射线晶体学和 NMR 进行了表征,而复合物已通过冷冻 EM 进行了表征。 没有完整的人类 eIF3 结构可用,但已在 43S PIC 的背景下以中等分辨率确定了近乎完整的复合体。 哺乳动物 eIF3 的结构核心通常被描述为具有拟人化特征的五叶颗粒,主要由 PCI/MPN 八聚体组成。 PCI 域以蛋白酶体帽 (P)、COP9 信号体 (C) 和 eIF3 (I) 之间的结构相似性命名,而 MPN 域则以与 Mpr1-PadI N 末端域的结构相似性命名。
信令
编辑eIF3 作为通过 S6K1 和 mTOR/Raptor 的细胞信号转导的枢纽。 特别是,eIF3 在其非活性状态下与 S6K1 结合,激活的 mTOR/Raptor 与 eIF3 结合并磷酸化 S6K1 以促进其从 eIF3 中释放。 磷酸化的 S6K1 然后可以自由磷酸化其自身的许多靶标,包括 eIF4B,从而充当翻译控制机制。
亚基
编辑eIF3 亚基在复合体中以相等的化学计量存在,但 eIF3J 除外,它松散结合并且对几个物种的生存能力不是必需的。 亚基最初是按哺乳动物的分子量按字母顺序排列的(A 最高),但分子量的排列因物种而异。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229299/