
分泌
编辑分流是物质从一点到另一点的运动,例如细胞或腺体分泌的化学物质。 相反,排泄是从细胞或生物体中去除某些物质或废物。 细胞分泌的经典机制是通过质膜上称为孔隙体的分泌入口。 孔隙体是嵌在细胞膜中的永久性杯状脂蛋白结构,分泌性囊泡在此处短暂停靠并融合以从细胞中释放囊泡内内容物。
细菌物种中的分酸是指效应分子的运输或易位,例如:蛋白质、酶或毒素(例如病原菌中的霍乱毒素,例如霍乱弧菌)从细菌细胞的内部(细胞质或细胞质)到外部。 分流是细菌在其自然周围环境中为适应和生存而发挥功能和运作的一种非常重要的机制。
真核细胞
编辑机制
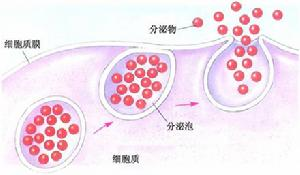
编辑包括人类细胞在内的真核细胞具有高度进化的分泌过程。 针对外部的蛋白质由停靠在粗面内质网 (ER) 上的核糖体合成。 当它们被合成时,这些蛋白质易位到内质网腔,在那里它们被糖基化,分子伴侣帮助蛋白质折叠。 错误折叠的蛋白质通常在这里被识别,并通过 ER 相关的降解逆向转移到胞质溶胶,在那里它们被蛋白酶体降解。 含有正确折叠蛋白质的囊泡随后进入高尔基体。
在高尔基体中,蛋白质的糖基化被修饰,并且可能发生进一步的翻译后修饰,包括切割和功能化。 然后蛋白质被移动到分泌囊泡中,分泌囊泡沿着细胞骨架移动到细胞边缘。 更多的修饰可以发生在分泌小泡中(例如,胰岛素从分泌小泡中的胰岛素原裂解)。
最终,通过称为胞吐作用的过程,囊泡在孔隙体处与细胞膜融合,将其内容物倾倒出细胞环境。
通过使用 pH 梯度对该序列进行严格的生化控制:胞质溶胶的 pH 值为 7.4,ER 的 pH 值为 7.0,顺式高尔基体的 pH 值为 6.5。 分泌囊泡的 pH 值介于 5.0 和 6.0 之间; 一些分泌囊泡演变成溶酶体,其 pH 值为 4.8。
非经典分泌物
许多蛋白质如 FGF1 (aFGF)、FGF2 (bFGF)、白介素-1 (IL1) 等没有信号序列。 他们不使用经典的 ER-GoLGi 通路。 这些是通过各种非经典途径分泌的。
至少描述了四种非经典(非常规)蛋白质分泌途径。 他们包括:
- 可能通过膜转运蛋白直接跨质膜转运蛋白质
- 起泡
- 溶酶体分泌
- 通过源自多泡体的外泌体释放
此外,蛋白质可以通过机械或生理损伤从细胞中释放出来,并通过用无血清培养基或缓冲液洗涤细胞诱导的质膜中的非致命性瞬时渗透孔释放。
在人体组织中
编辑许多人类细胞类型都具有成为分泌细胞的能力。 它们有发达的内质网和实现这一功能的高尔基体。 产生分泌物的组织包括分泌消化酶和胃酸的胃肠道,分泌表面活性剂的肺,以及分泌皮脂润滑皮肤和毛发的皮脂腺。 眼睑中的睑板腺分泌睑脂以润滑和保护眼睛。
在革兰氏阴性菌中
编辑分流并不是真核生物独有的——它也存在于细菌和古细菌中。 ATP 结合盒 (ABC) 型转运蛋白在生命的三个领域中很常见。 一些分泌蛋白通过 SecYEG 转运子跨细胞质膜转运,SecYEG 转运子是两个转运系统之一,它需要分泌蛋白上存在 N 末端信号肽。
其他通过双精氨酸易位途径 (Tat) 跨细胞质膜易位。 革兰氏阴性菌有两层膜,因此分泌物的拓扑结构更加复杂。 革兰氏阴性菌至少有六种专门的分泌系统。 许多分泌蛋白在细菌发病机制中特别重要。
I 型分泌系统(T1SS 或 TOSS)
编辑I 型分泌是一种伴侣依赖性分泌系统,使用 Hly 和 Tol 基因簇。 该过程开始于要分泌的蛋白质上的前导序列被 HlyA 识别并结合膜上的 HlyB。 这个信号序列对于 ABC 转运蛋白来说是极其特异的。 HlyAB 复合物刺激 HlyD,HlyD 开始展开并到达 TolC 识别的外膜。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/229493/