
白色念珠菌
编辑白色念珠菌是一种机会致病酵母,是人类肠道菌群的常见成员。它也可以在人体外生存。在40-60%的健康成人的胃肠道和口腔中检测到它。它通常是一种共生生物,但在各种条件下,它会在免疫功能低下的个体中致病。它是导致人类感染念珠菌病的少数念珠菌属物种之一,这是真菌过度生长的结果。例如,念珠菌病经常在HIV感染患者中观察到。白色念珠菌是从在(永久)植入医疗器械或人体组织上形成的生物膜中分离出来的最常见的真菌物种。白色念珠菌、热带念珠菌、近平滑念珠菌和光滑念珠菌共同导致50-90%的人类念珠菌病病例。据报道,由于白色念珠菌引起的全身性念珠菌病患者的死亡率为40%。
白色念珠菌通常用作真菌病原体的模式生物。它通常被称为二态真菌,因为它既可以作为酵母也可以作为丝状细胞生长。然而,它有几种不同的形态表型,包括不透明、GUT和假菌丝形式。白色念珠菌长期以来被认为是一种没有单倍体阶段的专性二倍体生物。然而,事实并非如此。除了单倍体阶段,白色念珠菌还可以存在于四倍体阶段。后者是在二倍体白色念珠菌时形成的当细胞处于不透明形式时,它们会交配。二倍体基因组大小约为29Mb,多达70%的蛋白质编码基因尚未表征。白色念珠菌很容易在实验室培养,可以在体内和体外进行研究。根据媒体的不同,可以进行不同的研究,因为媒体会影响白色念珠菌的形态状态。一种特殊类型的培养基是CHROMagar™念珠菌,可用于鉴定不同种类的念珠菌。
基因组
编辑白色念珠菌的基因组单倍体大小几乎为16Mb(二倍体阶段为28Mb),由8组染色体对组成,称为chr1A、chr2A、chr3A、chr4A、chr5A、chr6A、chr7A和chrRA。第二组(白色念珠菌是二倍体)具有相似的名称,但末尾有一个B。Chr1B、chr2B、…和chrRB。整个基因组包含6,198个开放阅读框(ORF)。这些ORF中有70%尚未表征。全基因组已被测序,使其成为最早被完全测序的真菌之一(仅次于酿酒酵母和粟酒裂殖酵母)。所有开放阅读框(ORF)也可在网关适应向量。除了这个ORFeome之外,还有一个GRACE(基因替换和条件表达)文库来研究白色念珠菌基因组中的必需基因。研究白色念珠菌最常用的菌株是WO-1和SC5314菌株。已知WO-1菌株以较高频率在白色不透明形式之间切换,而SC5314菌株是用于基因序列参考的菌株。
白色念珠菌基因组最重要的特征之一是高杂合性。这种杂合性的基础在于数字和结构染色体重排和变化的发生,作为通过染色体长度多态性(重复的收缩/扩展)、相互易位、染色体缺失、非同义单核苷酸多态性和个体的三体性产生遗传多样性的手段染色体。这些核型改变导致表型发生变化,这是一种适应这种真菌的策略。随着对白色念珠菌基因组的完整分析的可用性,这些机制正在进一步探索。
念珠菌属的一个不同寻常的特征是,在它的许多物种(包括白色念珠菌和热带念珠菌,但不包括光滑念珠菌)中,通常指定亮氨酸的CUG密码子在这些物种中指定丝氨酸。这是偏离标准遗传密码的一个不寻常的例子,大多数这样的偏离是在起始密码子中,或者对于真核生物来说,是线粒体遗传密码。在某些环境中,这种改变可能有助于这些念珠菌物种通过诱导永久性应激反应,一种更普遍的热休克反应形式。然而,这种不同的密码子使用使得研究模型生物酿酒酵母中的白色念珠菌蛋白质-蛋白质相互作用变得更加困难。为了克服这个问题,开发了一种白色念珠菌特异性双杂交系统。
白色念珠菌的基因组是高度动态的,由不同的CUG翻译贡献,并且这种变异性已被有利地用于该物种的分子流行病学研究和种群研究。基因组序列已允许识别白色念珠菌中是否存在副性循环(未检测到减数分裂)。这项关于六种念珠菌有性生殖进化的研究发现,最近主要减数分裂交叉形成途径的成分丢失,但保留了一条次要途径。作者建议,如果念珠菌物种经历减数分裂它是机械减少或不同的机械,并表明许多物种可能存在未被识别的减数分裂周期。在另一项进化研究中,将部分CUG身份重新定义(来自念珠菌属)引入酿酒酵母克隆引起了对有性生殖产生负面影响的应激反应。这种CUG身份重新定义发生在念珠菌物种的祖先中,被认为将这些物种锁定为二倍体或多倍体状态,可能会阻碍有性生殖。
白色念珠菌的形态
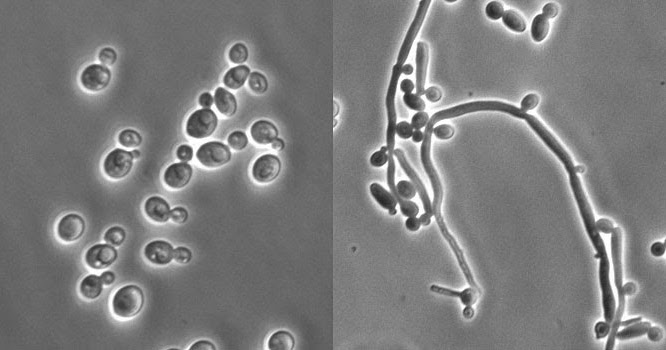
编辑由于表型转换和芽到菌丝的转变,白色念珠菌表现出广泛的形态表型。酵母到菌丝的转变(丝状化)是一个快速过程,由环境因素诱导。表型转换是自发的,以较低的速率发生,并且在某些菌株中,已知多达七种不同的表型。研究得最好的转换机制是白色到不透明的转换(表观遗传过程)。还描述了其他系统。DavidR.Soll及其同事发现了两个系统(高频开关系统和白色到不透明开关)。白色念珠菌的转换经常但不总是受环境条件的影响,例如CO2水平、厌氧条件、使用的介质和温度。在其酵母形式中,白色念珠菌的范围从10到12微米。孢子可以在称为厚生孢子的假菌丝上形成,当置于干燥或炎热季节等不利条件下时,它们仍能存活。
酵母到菌丝的转换
编辑尽管通常被称为二态性,但实际上白色念珠菌是多态性的(通常也称为多态性)。当在标准酵母实验室培养基中培养时,白色念珠菌生长为卵形“酵母”细胞。然而,温度、CO2、营养物质和pH值的温和环境变化可导致形态转变为丝状生长。丝状细胞与酵母细胞有许多相似之处。这两种细胞类型似乎都在白色念珠菌的存活和致病性中发挥了特定的、独特的作用.酵母细胞似乎更适合在血流中传播,而菌丝细胞被认为是一种毒力因子。菌丝细胞是侵入性的,据推测对于组织穿透、器官定植和存活以及逃逸的巨噬细胞很重要。从酵母到菌丝细胞的转变被称为白色念珠菌毒力的关键因素之一。但是,认为没有必要。当白色念珠菌细胞在模拟人类宿主生理环境的培养基中生长时,它们会生长为丝状细胞(真菌丝和假菌丝)。白色念珠菌也可以形成厚壁孢子,其功能仍然未知,但据推测它们在恶劣环境中发挥作用,因为它们通常是在不利条件下形成的。
圆形、白色相和细长、不透明相白色念珠菌细胞:比例尺为5µm在这个调节白色不透明开关的遗传网络模型中,白色和金色盒子分别代表富含白色和不透明状态的基因。蓝线代表基于遗传上位性的关系。红线代表每个基因的Wor1控制,基于染色质免疫沉淀实验中的Wor1富集。根据每个基因的白色和不透明状态表达推断激活(箭头)和抑制(条形)。
高频开关
编辑除了经过充分研究的酵母到菌丝的转变之外,还描述了其他转换系统。一种这样的系统是“高频开关”系统。在这种转换过程中,会自发产生不同的细胞形态(表型)。这种类型的切换不会大量发生,而是代表一个可变系统,并且它独立于环境条件发生。菌株3153A产生至少七种不同的菌落形态。在许多菌株中,不同的相在低频下自发地转换为其他相。这种转换是可逆的,菌落类型可以从一代遗传到另一代。能够通过如此多的不同(形态)表型进行转换,使得白色念珠菌能够在不同的环境中生长,既可以作为共生体,也可以作为病原体。
在3153A菌株中,已经发现了一种称为SIR2(沉默信息调节剂)的基因,该基因似乎对表型转换很重要。SIR2最初是在酿酒酵母(啤酒酵母)中发现的,它参与染色体沉默——一种转录调控形式,其中基因组的区域被染色质结构的变化可逆地灭活(染色质是构成染色体的DNA和蛋白质的复合物)。在酵母中,在这些沉默区域中发现了参与控制交配类型的基因,而SIR2通过在该区域维持沉默的染色质结构来抑制它们的表达。与表型转换有关的白色念珠菌SIR2的发现表明它也具有由SIR2控制的沉默区域,表型特异性基因可能存在于其中。SIR2本身是如何在酿酒酵母中被调控的,这可能会提供更多关于白色念珠菌转换机制的线索。
白色到不透明的切换
编辑在二态性和第一个描述的高频切换系统之后,白色念珠菌经历了另一个称为白色到不透明切换的高频切换过程,这是白色念珠菌中的另一个表型切换过程。这是在白色念珠菌中发现的第二个高频开关系统。白色到不透明的转换是一种表观遗传转换系统。表型转换通常用于指代白色不透明转换,它由两个阶段组成:一个阶段以圆形细胞的形式生长,呈光滑的白色菌落(称为白色形式),另一个阶段呈棒状并生长为扁平、灰色菌落(称为不透明形式)。这种从白细胞到不透明细胞的转变对于白色念珠菌的毒力和交配过程很重要,因为不透明形式是具有交配能力的形式,与白色类型相比,交配效率高一百万倍。白色和不透明形式之间的这种切换由WOR1调节器(白色到不透明调节器1)调节,该调节器由配对类型控制基因座(MTL)阻遏物(a1-α2)抑制WOR1的表达。除了白色和不透明相外,还有第三种:灰色表型。这种表型显示出引起皮肤感染的最高能力。白色、不透明和灰色表型形成三稳态表型转换系统。由于通常难以区分白色、不透明和灰色细胞,因此可以在培养基中添加一种染料phloxineB。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/134360/