
合成离子通道
编辑合成离子通道是从头进入脂质双层的化合物,形成孔隙,并允许离子从一侧流到另一侧。它们是天然离子通道的人造类似物,因此也称为人工离子通道。与生物通道相比,它们通常允许类似大小的通量,但
与天然通道一样,合成通道通常以单分子(例如,平面双层的电压钳)和集成技术(囊泡中的通量)的组合为特征。合成离子通道的研究可能会导致新的单分子传感技术以及新的治疗方法。
合成离子通道的历史
编辑虽然半合成离子通道(通常基于短杆菌肽等修饰肽通道)自1970年代以来已经制备,但在1982年首次尝试使用取代的β-环糊精制备合成离子通道。受短杆菌肽的启发,这种分子被设计成一个桶形实体,跨越双层膜的单个小叶,只有当相反小叶中的两个分子以端到端的方式聚集在一起时才会变得活跃。虽然该化合物确实在囊泡中诱导离子流,但数据并未明确显示通道形成(与其他传输机制相反;参见机制)。1989年至1990年,两组研究人员首次报道了通过这种通道进行的Na+转运。随着1990年代初期将电压钳技术应用于合成通道研究,研究人员能够观察到合成分子的量子化电活动,这通常被认为是离子通道的标志性证据。这导致了未来二十年研究活动的持续增加。2009年,发表了超过25篇关于该主题的同行评审论文,并提供了一系列综合评论。
表征和机制
编辑离子通过膜的被动传输可以通过三种主要机制进行:通过运输、通过破坏膜中的缺陷或通过定义的轨迹;这些对应于离子载体、去污剂和离子通道转运蛋白。虽然合成离子通道研究试图制备通过特定路径显示电导的化合物,但阐明机理很困难,而且很少明确。两种主要的表征方法都有其缺点,因此,通常定义了功能但假定了机制。
合奏,基于囊泡的时间课程
编辑离子传输的一条证据来自对统计系综的宏观检查。所有这些技术都使用具有截留体积的完整囊泡,并通过不同的光谱方法报告离子通道活动。在典型情况下,染料被包裹在囊泡群体中。选择该染料以比色或荧光响应离子的存在;这种离子通常不存在于囊泡内部,但存在于外部。如果没有离子转运蛋白,脂质双层作为阻止离子通量的动力屏障,染料会无限期地保持黑暗。由于离子转运体允许外部的离子扩散进来,它的添加会影响染料的颜色/荧光特性。通过随时间宏观监测染料的性质,并控制外部因素,可以测量化合物作为离子转运体的能力。然而,观察离子传输并不能确定离子通道是机制。任何类型的转运蛋白都可能导致相同的观察结果,并且通常需要额外的确凿证据。旨在探测选择性、门控和其他通道参数的复杂实验在过去的二十年中得到了发展,最近得到了总结。
囊泡测定变化
随机的、基于平面双层的电流轨迹
编辑上述基于集合的方法的替代方法是电压钳实验。在电压钳实验中,两个电解液隔室被一个孔径分开,孔径通常在5-250微米之间。脂质双层涂在这个孔上,从而将隔室电分离;分子性质可以通过测量其电容来确定。在添加(理想)离子通道后,在两个隔室之间形成了明确的路径。通过该孔,离子迅速沿电位和电化学梯度(>106/秒)向下流动,最大通量受孔的几何形状和尺寸限制。在稍后的某个时刻,孔隙可能会关闭或塌陷,因此电流会回到零。这种由单分子事件产生和放大的开态电流通常在pA到nA的数量级上,时间分辨率约为。毫秒。理想或接近理想的事件在文献中被称为方顶,并且被认为是基于通道的机制的签名。值得注意的是,在这个尺度上观察到的事件是真正随机的——也就是说,它们是随机分子碰撞和构象变化的结果。由于膜面积远大于孔,因此多个副本可以相互独立地打开和关闭,从而产生阶梯状外观(图中的面板C);这些理想事件通常被建模为马尔可夫过程。通过使用活动网格符号,对1982-2010年期间用电压钳法研究的合成离子通道进行了批判性审查。虽然理想轨迹在文献中最常被分析和报告,但许多记录显然是不理想的,其中一个子集被证明是分形的。开发分析这些非理想痕迹并阐明它们与传输机制的关系的方法是当代研究的一个领域。
合成离子通道例子
编辑据报道,大量的合成分子在脂质膜中充当离子转运体。此处描述了一个选择,以展示可行结构和可实现功能的广度。截至2010年的文献综合评论可在三方系列中获得。
按化学结构
编辑大多数(但不是全部;参见极简通道)合成通道的化学结构远大于典型的小分子(分子量约1-5kDa)。这源于对两亲性的需求,即具有足够的疏水部分以允许分配到脂质双层中,以及极性或带电的头部基团以相对于膜确定确定的方向和几何形状。
基于大环
基于冠醚
杯芳烃基
含有环尺寸为3和4的杯芳烃的离子通道均有报道。对于杯[4]芳烃,有两种构象可供使用,并且已经开发了1,3-alt和锥体构象的例子。
基于环糊精
第一个合成离子通道是通过对β-环糊精初级边缘的部分取代而构建的。此后已报道了其他取代的β-环糊精,包括硫醇改性的环糊精、阴离子选择性低聚丁烯通道和各种聚环氧乙烷连接的星爆低聚物。最近报道了通过点击化学制备的大量环糊精半通道的构效关系。
刚性棒
基于肽
已知交替的D/L肽大环可自聚集成纳米管,并且已证明所得纳米管可充当脂质膜中的离子通道。其他架构使用肽螺旋作为支架来连接其他功能,例如不同大小的冠醚。这些肽冠通道的特性很大程度上取决于封端基团的特性。
极简主义频道
各种各样的
基于G-四重奏的频道
金属有机通道
混合生物通道
构建了通过修饰天然离子通道构建的半合成生物杂化通道。利用现代合成有机化学,这些允许对现有结构进行精确修改,以阐明其传输机制或嫁接新功能。Gramicidin和alamethicin一直是选择性修饰的流行起点。上图说明了一个示例,其中冠醚固定在离子通过入口的口部。在电压钳实验中,该通道显示离散电导,但离子选择性与野生型短杆菌肽不同。虽然使用诱变来修饰大蛋白质通道通常被认为超出了合成通道的范围,但界限并不清晰,正如环糊精与α-溶血素的超分子或共价键合所证明的那样。
按运输特点
编辑离子通道可以通过其开放特性、离子选择性和通量控制(门控)来表征。许多合成离子通道在这些方面的一个或多个方面显示出独特的特性。
开口特点
离子通道形成分子通常可以在平面双层膜中显示多种类型的电导活动。这些行动模式中的每一种都可以通过其
这些事件在其整个持续时间中不一定是一致的,因此各种形状的导电迹线都是可能的。
离子选择性
大多数合成离子通道对碱金属阳离子的选择性遵循EisenmanI序列(Cs+>Rb+>K+>Na+>>Li+),这表明选择性的来源取决于从水中去除水所需的能量差异完全水合的阳离子。一些合成通道显示出其他离子选择性模式,并且仅报告了遵循相反选择性序列(EisenmanXI;Cs+<Rb+<K+<Na+<<Li+)的合成通道的单一实例。
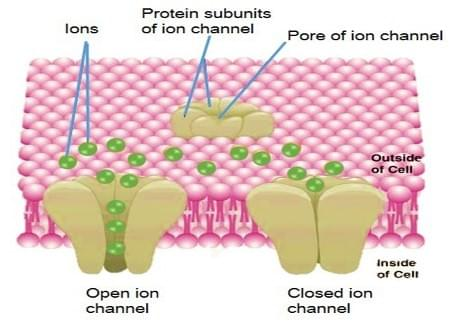
门控
电压响应
大多数合成通道的电导是欧姆的,也就是说,通过的电流(无论是单独的还是整体的)与跨膜的电位成正比。然而,一些罕见的通道显示出非线性的电流-电压特性。具体来说,已知两种不同类型的非欧姆电导:
- 一种整流行为,其中电流通过取决于所施加电位的符号,以及
- 指数电位依赖性,其中通过的电流与施加的电位成指数关系。
前者要求脂质双层的中平面不对称,并且通常通过引入整体分子偶极子来实现。后者在天然通道(如alamethicin)中得到证明,在合成离子通道中很少遇到。它们可能与脂质离子通道有关,但迄今为止它们的机制仍然难以捉摸。
配体反应
某些合成离子通道具有可以通过额外的外部化学物质调节的电导。上调(通道被配体打开)和下调(通道被配体关闭)都是已知的:不同的机制,包括超分子聚集体的形成,以及分子间和分子内的阻塞。
其他
对其他信号作出反应的监管要素是已知的;例子包括光调制电导以及通过氨基甲酸酯基团的异构化构造的热开关。迄今为止,尚未报道机械敏感的合成离子通道。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/151226/