
纤维素酶
编辑纤维素酶是主要由真菌、细菌和原生动物产生的几种酶中的任何一种,可催化纤维素分解、纤维素和一些相关多糖的分解。该名称也用于任何天然存在的混合物或各种此类酶的复合物,它们连续或协同作用以分解纤维素材料。
纤维素酶将纤维素分子分解成单糖(“单糖”),例如β-葡萄糖,或更短的多糖和寡糖。纤维素分解具有相当大的经济意义,因为它使植物的主要成分可用于消费和用于化学反应。所涉及的特定反应是水解的1,4-β-D-的糖苷键的纤维素,半纤维素,地衣淀粉和谷类的β-d葡聚糖。由于纤维素分子相互之间结合牢固,与其他多糖(如淀粉)的分解相比,纤维素分解相对困难。
大多数哺乳动物自身消化纤维素等膳食纤维的能力非常有限。在许多食草动物(如牛和羊等反刍动物和马等后肠发酵罐)中,纤维素酶是由共生细菌产生的。内源性纤维素酶由几种类型的后生动物产生,例如一些白蚁、蜗牛、和蚯蚓。
近日,纤维素酶也已经在绿色微藻(发现莱茵衣藻,Goniumpectorale和团藻carteri)和属于他们的催化结构域(CD)GH9家庭显示最高序列同源性后生动物内源性纤维素酶。藻类纤维素酶是模块化的,除了一些成员中假定的Ig样和未知结构域外,还包括假定的新型富含半胱氨酸的碳水化合物结合模块(CBM)、富含脯氨酸/丝氨酸(PS)的接头。来自胸甲藻的纤维素酶由两个由接头隔开的CD和一个C端CBM组成。
几种不同种类的纤维素酶是已知的,它们在结构和机理上不同。与“纤维素酶”名称相关的同义词、衍生物和特定酶包括内切-1,4-β-D-葡聚糖酶(β-1,4-葡聚糖酶、β-1,4-内切葡聚糖水解酶、内切葡聚糖酶D、1,4-(1,3,1,4)-β-d葡聚糖4-葡聚糖水解酶),羧甲基纤维素(CMC酶),微晶纤维素酶,纤维糊精酶,纤维素酶甲,木粉AP,碱纤维素酶,纤维素酶A3,9.5纤维素酶,和pancellaseSS.裂解木质素的酶偶尔被称为纤维素酶,但这种旧用法已被弃用;它们是木质素修饰酶。
纤维素酶的类型和动作
编辑基于催化反应类型的五种一般纤维素酶类型:
- 内切纤维素酶(EC3.2.1.4)在无定形位点随机切割内部键,从而产生新的链端。
- 外切纤维素酶或纤维二糖水解酶(EC3.2.1.91)从内切纤维素酶产生的暴露链的末端切割2到4个单元,产生四糖或二糖,例如纤维二糖。外切纤维素酶进一步分为I型,从纤维素链的还原端开始进行性工作,以及II型,从非还原端开始进行性工作。
- 纤维二糖酶(EC3.2.1.21)或β-葡萄糖苷酶将外切纤维素酶产物水解成单个单糖。
- 氧化纤维素酶通过自由基反应解聚纤维素,例如纤维二糖脱氢酶(受体)。
- 纤维素磷酸化酶使用磷酸盐而不是水来解聚纤维素。
微晶纤维素酶几乎完全具有外切纤维素酶活性,因为微晶纤维素是一种高度微晶的底物。
在上述类型中,还有渐进式(也称为进行式)和非渐进式类型。渐进式纤维素酶将继续与单条多糖链相互作用,非渐进式纤维素酶将相互作用一次,然后脱离并接合另一条多糖链。
纤维素酶的作用被认为是协同的,因为所有三类纤维素酶比单独添加所有三类纤维素酶能产生更多的糖。除了反刍动物,大多数动物(包括人类)在体内不产生纤维素酶,只能通过发酵部分分解纤维素,限制了它们在纤维植物材料中使用能量的能力。
纤维素酶的结构
编辑大多数真菌纤维素酶具有双结构域结构,具有一个催化结构域和一个纤维素结合结构域,它们通过柔性接头连接。这种结构适用于在不溶性底物上工作,它允许酶以类似毛毛虫的方式在表面上二维扩散。然而,也有缺乏纤维素结合域的纤维素酶(主要是内切葡聚糖酶)。
底物的结合和催化作用都依赖于酶的三维结构,这是蛋白质折叠水平的结果。发生在活性位点内的氨基酸序列和残基排列、底物结合的位置,可能会影响配体的结合亲和力、活性位点内底物的稳定性和催化作用等因素。底物结构与酶的精确活性位点结构互补。残基位置的变化可能会导致这些相互作用中的一种或多种发生扭曲。温度、pH值和金属离子等其他因素会影响酶结构之间的非共价相互作用。Thermotogamaritima物种产生的纤维素酶由2个β-折叠(蛋白质结构)组成,围绕一个中心催化区域,该区域是活性位点。该酶被归类为内切葡聚糖酶,它在内部裂解纤维素链中的β-1,4-糖苷键,促进聚合物的进一步降解。与T.Maritima同属的不同物种产生具有不同结构的纤维素酶。由CoprinopsisCinerea物种产生的纤维素酶由七个蛋白质链组成,呈封闭隧道形状,称为β/α桶。这些酶水解底物羧甲基纤维素。底物与活性位点的结合会引起构象的变化,从而使分子降解。
纤维素酶复合物
编辑在许多细菌中,体内的纤维素酶是在超分子复合物(纤维素小体)中组织起来的复杂酶结构。它们可以包含但不限于五种不同的酶亚基,代表即内切纤维素酶、外切纤维素酶、纤维二糖酶、氧化纤维素酶和纤维素磷酸化酶,其中只有外切纤维素酶和纤维二糖酶参与β(1→4)键的实际水解。组成纤维素体的亚单位的数量也可以决定酶活性的速率。
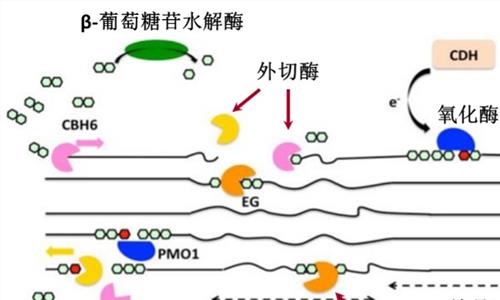
多域纤维素酶在许多分类群中广泛存在,然而,在纤维素体中发现的来自厌氧细菌的纤维素酶具有最复杂的结构,由不同类型的模块组成。例如,ClostridiumcellulolytICUm产生13种GH9模块化纤维素酶,其中包含不同数量和排列的催化域(CD)、碳水化合物结合模块(CBM)、dockerin、接头和Ig样域。
例如,来自里氏木霉的纤维素酶复合物包含标记为C1的组分(57,000道尔顿),其分离结晶纤维素链、内切葡聚糖酶(约52,000道尔顿)、外切葡聚糖酶(约61,000道尔顿)和β-葡糖苷酶(约61,000道尔顿)道尔顿)。
在产生纤维素体的细菌基因组中已经鉴定出许多被称为dockerins和cohesins的“特征”序列。根据它们的氨基酸序列和三级结构,纤维素酶分为氏族和科。
由于酶与纤维素底物之间的紧密接近,多模块纤维素酶比游离酶(只有CD)更有效,这是由于协同作用。CBM参与纤维素的结合,而糖基化接头为CD提供灵活性,以提高活性和蛋白酶保护,并增加与纤维素表面的结合。
纤维素酶的用途
编辑纤维素酶用于咖啡的商业食品加工。它在豆类干燥过程中进行纤维素的水解。此外,纤维素酶广泛用于纺织工业和洗衣洗涤剂。它们还用于纸浆和造纸工业的各种用途,甚至用于制药应用。纤维素酶用于将生物质发酵成生物燃料,尽管目前该过程相对处于实验阶段。在医学上,纤维素酶用于治疗植物牛黄,这是一种在人胃中发现的纤维素牛黄,并且它通过水解细胞外聚合物(EPS)的结构基质胞外多糖中的β(1-4)糖苷键,在降解多微生物细菌生物膜方面表现出功效。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/129120/