简介
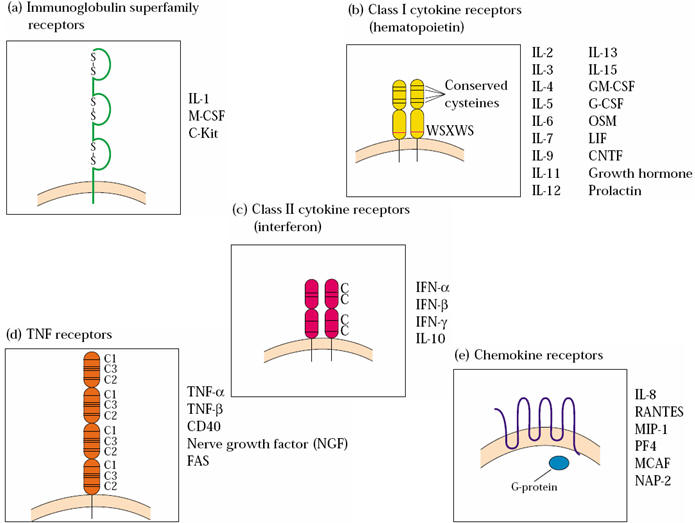
编辑趋化因子受体是在某些细胞表面发现的细胞因子受体,这些细胞因子与一种称为趋化因子的细胞因子相互作用。 在人类中发现了 20 种不同的趋化因子受体。 每一种都具有视紫红质样 7 跨膜 (7TM) 结构,并与 G 蛋白偶联以在细胞内进行信号转导,从而使它们成为 G 蛋白偶联受体的一个大蛋白家族的成员。 在与其特定的趋化因子配体相互作用后,趋化因子受体触发细胞内钙 (Ca2+) 离子的流动(钙信号)。 这会引起细胞反应,包括称为趋化性的过程的开始,该过程将细胞运送到生物体内的所需位置。
趋化因子受体分为不同的家族,CXC趋化因子受体、CC趋化因子受体、CX3C趋化因子受体和XC趋化因子受体对应于它们结合的4个不同的趋化因子亚家族。 四个趋化因子受体家族在受体 N 末端附近的半胱氨酸残基间距不同。
结构特点
编辑趋化因子受体是含有 7 个跨膜结构域的 G 蛋白偶联受体,主要存在于白细胞表面,使其成为视紫红质样受体之一。 迄今为止,已经鉴定了大约 19 种不同的趋化因子受体,它们具有许多共同的结构特征; 它们由大约 350 个氨基酸组成,分为一个短而酸性的 N 末端、七个具有三个细胞内亲水环和三个细胞外亲水环的螺旋跨膜结构域,以及一个含有充当磷酸化位点的丝氨酸和苏氨酸残基的细胞内 C 末端 在受体调节过程中。 趋化因子受体的前两个细胞外环通过两个保守的半胱氨酸残基之间的二硫键连接在一起。 趋化因子受体的 N 末端与趋化因子结合,对配体特异性很重要。 G 蛋白与 C 末端偶联,这对于配体结合后的受体信号传导很重要。 尽管趋化因子受体在其一级序列中具有高度氨基酸同一性,但它们通常结合有限数量的配体。 趋化因子受体的功能是多余的,因为不止一种趋化因子能够与单个受体结合。
信号转导
编辑趋化因子受体的细胞内信号依赖于邻近的 G 蛋白。 G蛋白以异源三聚体形式存在; 它们由三个不同的亚基组成。 当分子 GDP 与 G 蛋白亚基结合时,G 蛋白处于非活性状态。 在趋化因子配体结合后,趋化因子受体与 G 蛋白结合,允许 GDP 交换另一种称为 GTP 的分子,以及不同 G 蛋白亚基的解离。 称为 Gα 的亚基可激活与细胞膜相关的称为磷脂酶 C (PLC) 的酶。 PLC 裂解磷脂酰肌醇 (4,5)-二磷酸 (PIP2) 形成两个第二信使分子,称为肌醇三磷酸 (IP3) 和甘油二酯 (DAG); DAG 激活另一种称为蛋白激酶 C (PKC) 的酶,而 IP3 触发细胞内储存的钙释放。 这些事件促进许多信号级联,影响细胞反应。
例如,当 CXCL8 (IL-8) 与其特定受体 CXCR1 或 CXCR2 结合时,细胞内钙的升高会激活磷脂酶 D (PLD),进而启动称为 MAP 激酶通路的细胞内信号级联。 同时,G蛋白亚基Gα直接激活一种称为蛋白酪氨酸激酶(PTK)的酶,使趋化因子受体尾部的丝氨酸和苏氨酸残基磷酸化,导致其脱敏或失活。 启动的 MAP 激酶通路激活特定的细胞机制,这些机制涉及趋化性、脱颗粒、超氧阴离子的释放,以及称为整合素的细胞粘附分子亲和力的变化。 趋化因子及其受体在癌症转移中起着至关重要的作用,因为它们参与外渗、迁移、微转移和血管生成。 趋化因子的这种作用与它们将白细胞定位到炎症部位的正常功能惊人地相似。

趋化因子受体 5 (CCR5) 的选择压力
编辑人类免疫缺陷病毒使用 CCR5 受体来靶向和感染人类的宿主 T 细胞。 它通过破坏 CD4+ T 辅助细胞削弱免疫系统,使身体更容易受到其他感染。 CCR5-Δ32 是 CCR5 基因的等位基因变体,具有 32 个碱基对缺失,导致截短的受体。 具有这种等位基因的人对艾滋病有抵抗力,因为艾滋病毒不能与非功能性 CCR5 受体结合。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/228843/