
植物抗病性
编辑植物抗病性通过两种方式保护植物免受病原体侵害:通过预先形成的结构和化学物质,以及通过感染引起的免疫系统反应。相对于易感植物,抗病性是指植物上或植物中病原体的生长减少,而术语“ 疾病耐受性”表示尽管病原体水平很高,但几乎没有病害的植物。疾病的结果取决于病原体,植物和环境条件的三方面相互作用(一种称为疾病三角形的相互作用)。
防御激活化合物可以在植物的血管系统中在细胞之间移动并系统地移动。但是,植物没有循环免疫细胞,因此大多数细胞类型都具有广泛的抗微生物防御能力。尽管在比较多个标本时可以观察到明显的抗病性质变(允许在相同环境中以相同接种量感染同一病原体菌株后分类为“抗性”或“易感性”),但是抗病性的定量差异是由通常在植物株或基因型之间观察到。植物始终抵抗某些病原体,但会屈服于其他病原体。抗药性通常是特定于某些病原体种类或病原体菌株的。
常见的抗病机制
编辑预制结构和化合物
编辑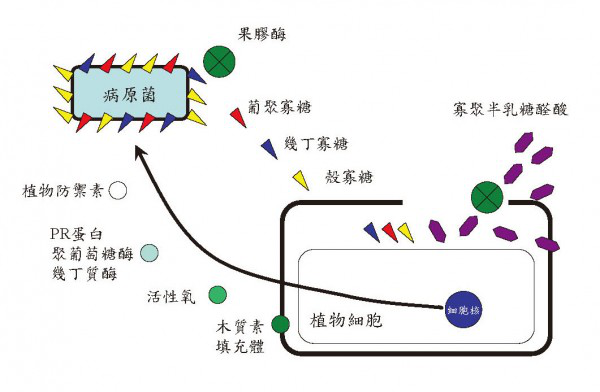
诱导型感染后植物防御
编辑- 细胞壁增强(纤维素、木质素、木栓质、细胞壁蛋白)
- 抗菌化学物质,包括活性氧,例如过氧化氢或过氧亚硝酸盐,或更复杂的植物抗毒素,例如染料木黄酮或卡马新
- 抗菌蛋白,例如防御素、硫蛋白或PR-1
- 抗菌酶,例如几丁质酶、β- 葡聚糖酶或过氧化物酶
- 超敏反应 -由“抵抗基因”介导的与防御相关的快速宿主细胞死亡反应。
免疫系统
编辑植物免疫系统带有两层相互连接的受体,一层最常感测细胞外的分子,另一层最常感测细胞内的分子。两种系统都可以感知入侵者并通过激活被感染细胞和邻近细胞中的抗菌防御来做出反应。在某些情况下,防御激活信号会传播到植物的其余部分甚至邻近的植物。这两个系统检测不同类型的病原体分子和植物受体蛋白的类别。
第一层主要由模式识别受体控制,该模式识别受体通过识别进化上保守的病原体或微生物相关分子模式(PAMP或MAMP)来激活。PRR的激活导致细胞内信号传导,转录重编程以及限制殖民化的复杂输出反应的生物合成。该系统称为PAMP触发免疫或模式触发免疫(PTI)。
第二层主要由R基因产物控制,通常被称为效应子触发免疫(ETI)。ETI通常通过特定病原体“效应物”的存在而被激活,然后触发强烈的抗微生物反应。
除PTI和ETI外,还可以通过感测与损害相关的化合物(DAMP)(例如在病原体感染过程中释放的部分植物细胞壁)来激活植物防御。
由PTI和ETI受体激活的反应包括离子通道门控、氧化爆发、细胞氧化还原变化或蛋白激酶级联反应,它们直接激活细胞变化(例如细胞壁增强或抗菌素产生),或激活基因表达变化,进而增强其他防御能力回应
植物免疫系统表现出与一些机械相似的免疫系统,昆虫和哺乳动物的,但也表现出许多植物特有的特征。上述两个层次对于植物免疫至关重要,但并未完全描述植物免疫系统。另外,许多明显的PTI或ETI的特定示例违反了通用的PTI / ETI定义,这表明需要扩大定义和/或范式。
模式触发的抗扰性
编辑PAMPs是居住在多种病原体属中的保守分子,被许多研究人员称为MAMPs。MAMP感知诱导的防御足以排斥大多数病原体。但是,病原体效应蛋白适用于抑制基础防御,例如PTI。已发现许多MAMP(和DAMP)受体。MAMP和DAMP通常通过携带LRR或LysM细胞外结构域的跨膜受体激酶来检测。
效应器触发的免疫力
编辑效应物触发的免疫(ETI)被病原体效应物激活。ETI反应依赖于R基因,并由特定的病原体菌株激活。植物ETI常常引起细胞凋亡的 超敏反应。
植物育种抗病
编辑植物育种者强调抗病植物系的选择和发展。植物病害也可以通过使用农药和耕作方法(例如轮作、耕作、种植密度、无病种子和设备清洁)进行部分控制,但是通常首选具有固有(遗传决定)抗病性的植物品种。抗病育种始于植物刚被驯化时。繁殖工作继续进行,因为病原体种群处于选择压力之下为了提高毒力,会出现新的病原体,不断发展的耕作方法和不断变化的气候会降低抗药性和/或增强病原体,而其他性状的植物育种也会破坏以前的抗药性。对一种病原体具有可接受抗性的植物品系可能缺乏对其他病原体的抗性。
抗性育种通常包括:
- 鉴定在其他方面可能不太理想但具有有用的抗病性状的植物,包括通常表现出增强抗性的野生植物品系。
- 将理想的但易感病的品种与作为抗病源的植物杂交。
- 在有利于疾病的环境中生长候选候选物,可能包括病原体接种。必须注意特定的病原体分离株,以解决单个病原体物种内的变异性。
- 保留其他理想特性(例如产量、品质和包括其他抗病特性)的抗病个体的选择。
如果随着病原体种群的发展,它在多年的广泛使用中继续有效,那么这种抗性就被称为持久性。“ 垂直抗性 ”特定于病原体物种的某些种族或品系,通常由单个R基因控制,并且耐久性较差。对整个病原体的水平或广谱抗药性通常仅是不完全有效,而是更持久,并且通常由在繁殖种群中分离的许多基因控制。
诸如马铃薯、苹果、香蕉和甘蔗等农作物通常通过无性繁殖来繁殖,以保留高度理想的植物品种,因为对于这些物种而言,异源杂交严重破坏了其优选性状。另见无性繁殖。通过植物转化的生物技术方法来管理影响疾病抗性的基因,无性繁殖作物可能是提高抗性的最佳目标之一。
科学的抗病育种起源于罗兰·比芬爵士(Sir RowlAND Biffen),他确定了单一的隐性基因来抵抗小麦黄锈病。然后,几乎每一种作物都经过育种以包含抗病(R)基因,其中许多是通过与野生亲缘种互渗获得的。
转基因或转基因工程抗病性
编辑术语GM(“基因修饰”)通常用作转基因的同义词,指使用重组DNA技术修饰的植物。具有转基因/转基因抗虫害能力的植物作为商业产品已经非常成功,尤其是在玉米和棉花中,并且每年在全球20多个国家的2000万公顷的土地上种植。1986年首次证明了转基因植物对微生物病原体的抗药性。病毒外壳蛋白基因序列的表达通过小RNA赋予病毒抗药性。事实证明,这是抑制病毒复制的广泛应用的机制。科学家结合了三种不同病毒的外壳蛋白基因,开发出了具有现场验证的多病毒抗性的南瓜杂交种。通过常规育种尚未达到对这种病毒的相似水平的抗性。
采取了类似的策略来对抗木瓜环斑病毒,该病毒到1994年威胁要摧毁夏威夷的木瓜产业。田间试验证明了出色的功效和高品质的水果。到1998年,第一批抗病毒的转基因木瓜被批准出售。抗病性已持续超过15年。转基因木瓜约占夏威夷产量的85%。
表达赋予对马铃薯卷叶病毒抗性的病毒复制酶序列的马铃薯品系以商品名NewLeaf Y和NewLeaf Plus出售,并在1999-2001年被商业生产广泛接受,直到麦当劳公司决定不购买转基因马铃薯,孟山都公司决定关闭他们的NatureMark马铃薯业务。NewLeaf Y和NewLeaf Plus马铃薯具有两个转基因特性,因为它们还表达了Bt介导的对科罗拉多马铃薯甲虫的抗性。
到2013年,没有其他对微生物病原体具有工程抗病性的农作物上市,尽管有十几种处于某种开发和测试状态。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/112678/