简介
编辑铁硫蛋白(或英式拼写中的铁硫蛋白)是一种蛋白质,其特征是存在铁硫簇,其中包含处于不同氧化态的硫化物连接的二、三和四铁中心。铁硫簇存在于多种金属蛋白中,例如铁氧还蛋白,以及 NADH 脱氢酶、氢化酶、辅酶 Q - 细胞色素 c 还原酶、琥珀酸 - 辅酶 Q 还原酶和固氮酶。 铁硫簇因其在线粒体和叶绿体中电子传递的氧化还原反应中的作用而广为人知。氧化磷酸化的复合物 I 和复合物 II 都具有多个 Fe-S 簇。它们具有许多其他功能,包括乌头酸酶所示的催化作用、SAM 依赖性酶所示的自由基生成以及硫辛酸和生物素生物合成中的硫供体。此外,一些 Fe-S 蛋白调节基因表达。 Fe-S 蛋白容易受到生物一氧化氮的攻击,形成二亚硝基铁络合物。在大多数 Fe-S 蛋白中,Fe 的末端配体是硫醇盐,但也有例外。
这些蛋白质在大多数生物体的代谢途径中普遍存在,导致一些科学家推测铁硫化合物在铁硫世界理论中对生命起源具有重要作用。
结构图案
编辑在几乎所有的 Fe-S 蛋白中,Fe 中心都是四面体,末端配体是来自半胱氨酰残基的硫醇硫中心。 硫化物基团是二配位或三配位的。 具有这些特征的三种不同类型的 Fe-S 团簇最为常见。
结构-功能原则
为了服务于它们的各种生物学作用,铁硫蛋白影响快速电子转移并跨越从 -600 mV 到 +460 mV 的整个生理氧化还原电势范围。
铁粉蛋白参与各种生物电子传输过程,例如光合作用和细胞呼吸,这些过程需要快速电子传输来维持生物体的能量或生化需求。
Fe3+-SR 键具有异常高的共价性,这是预期的。 将 Fe3+ 的共价性与 Fe2+ 的共价性进行比较时,Fe3+ 的共价性几乎是 Fe2+ 的两倍(20% 至 38.4%)。 Fe3+ 也比 Fe2+ 稳定得多。 像 Fe3+ 这样的硬离子通常具有低共价性,因为金属最低未占分子轨道与配体最高占分子轨道的能量不匹配。
靠近活性位点的蛋白质处存在来自外部 H2O 的 HO-H-S-Cys H 键,这种 H 键减少了从 Cys-S 供体到 Fe3+/2+ 的孤对电子捐赠。 使用冻干法去除这些外部 H2O 会导致 Fe-S 共价性增加,这意味着 H2O 正在降低共价性,因为 HOH-S 氢键会拉动硫电子。 由于共价键比 Fe2+ 更能稳定 Fe3+,因此 Fe3+ 更容易因 HOH-S 氢键而不稳定。
Fe3+ 3d 轨道能量遵循“反转”键合方案,偶然地使 Fe3+ d 轨道在能量上与硫 3p 轨道紧密匹配,从而在生成的键合分子轨道中提供高共价性。 这种高共价性降低了内球重组能,最终有助于快速电子转移。
2Fe–2S团簇
最简单的多金属系统,[Fe2S2] 簇,由两个铁离子和两个硫化物离子桥接并由四个半胱氨酰配体(在 Fe2S2 铁氧还蛋白中)或两个半胱氨酸和两个组氨酸(在 Rieske 蛋白中)配位构成。 氧化的蛋白质含有两个 Fe3+ 离子,而还原的蛋白质含有一个 Fe3+ 和一个 Fe2+ 离子。 这些物质以两种氧化态存在,(FeIII)2 和 FeIIIFeII。 CDGSH 铁硫结构域也与 2Fe-2S 簇有关。
4Fe–4S团簇
一个常见的主题是四个铁离子和四个硫离子位于立方烷型簇的顶点。 Fe 中心通常由半胱氨酰配体进一步配位。 [Fe4S4] 电子转移蛋白([Fe4S4] 铁氧还蛋白)可进一步细分为低电位(细菌型)和高电位(HiPIP)铁氧还蛋白。 低电位和高电位铁氧还蛋白通过以下氧化还原方案相关:
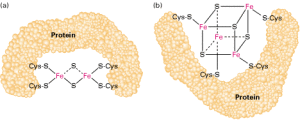

在 HiPIP 中,簇在 [2Fe3+, 2Fe2+] (Fe4S42+) 和 [3Fe3+, Fe2+] (Fe4S43+) 之间穿梭。 这种氧化还原对的电势范围为 0.4 至 0.1 V。在细菌铁氧还蛋白中,一对氧化态为 [Fe3+, 3Fe2+] (Fe4S4+) 和 [2Fe3+, 2Fe2+] (Fe4S42+)。 这种氧化还原对的电势范围为 -0.3 至 -0.7 V。4Fe-4S 簇的两个家族共享 Fe4S42+ 氧化态。 氧化还原对的差异归因于氢键的程度,这强烈地改变了半胱氨酰硫醇配体的碱性。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/215979/