
脂肪酸代谢
编辑脂肪酸代谢由涉及脂肪酸或与脂肪酸密切相关的各种代谢过程组成,脂肪酸是属于脂质常量营养素类别的分子家族。这些过程主要可分为(1)产生能量的分解代谢过程和(2)合成代谢过程,它们作为其他化合物的构建块。
在分解代谢中,脂肪酸被代谢以产生能量,主要以三磷酸腺苷(ATP)的形式存在。与其他常量营养素类别(碳水化合物和蛋白质)相比,当脂肪酸通过β氧化和柠檬酸循环完全氧化为CO2和水时,以每克能量为基础产生最多的ATP。因此,脂肪酸(主要以甘油三酯的形式)是大多数动物最重要的燃料储存形式,在植物中的程度较低。
在合成代谢中,完整的脂肪酸是甘油三酯、磷脂、第二信使、激素和酮体的重要前体。例如,磷脂形成磷脂双层,细胞的所有膜均由脂肪酸构成。磷脂包括质膜和其他包裹细胞内所有细胞器的膜,例如细胞核、线粒体、内质网和高尔基体。在另一种类型的合成代谢中,脂肪酸被修饰以形成无数其他化合物。例如,一些第二信使和局部激素。由储存在细胞膜中的花生四烯酸制成的前列腺素可能是这些局部激素中最著名的一组。
脂肪酸分解代谢
编辑血液中高肾上腺素和低胰岛素水平诱导的脂解过程(在脂肪细胞中)的示意图。肾上腺素与脂肪细胞细胞膜中的β-肾上腺素能受体结合,导致细胞内产生cAMP。cAMP激活蛋白激酶,该蛋白激酶磷酸化,从而激活脂肪细胞中的激素敏感性脂肪酶。这种脂肪酶将游离脂肪酸从它们与储存在脂肪细胞脂肪滴中的脂肪中的甘油的连接中分离出来。然后游离脂肪酸和甘油被释放到血液中。然而,最近的研究表明,脂肪甘油三酯脂肪酶必须首先将甘油三酯转化为甘油二酯,激素敏感性脂肪酶将甘油二酯转化为甘油单酯和游离脂肪酸。甘油单酯被甘油单酯脂肪酶水解。激素敏感性脂肪酶的活性受循环激素胰岛素、胰高血糖素、去甲肾上腺素和肾上腺素的调节,如图所示。的输送的示意图游离脂肪酸在附着于血液血浆白蛋白,其跨细胞膜使用蛋白转运扩散,并且其活化,使用ATP,以形成酰基-CoA在胞质溶胶中。出于图解目的,插图是12碳脂肪酸。人体血浆中的大多数脂肪酸长度为16或18个碳原子。肉碱-酰基-CoA转移酶(CAT)将酰基-CoA分子转移到线粒体内膜的示意图。出于图示目的,所示的酰基链只有12个碳原子长。人体血浆中的大多数脂肪酸长度为16或18个碳原子。细胞质中高浓度的丙二酰辅酶A(脂肪酸合成的第一步)会抑制CAT。这意味着脂肪酸合成和脂肪酸分解代谢不能在任何给定细胞中同时发生。线粒体基质中酰基辅酶A分子的β氧化过程的示意图。在这个过程中,形成了一个比过程开始时短2个碳原子的酰基辅酶A分子。乙酰辅酶A、水和5个ATP分子是每个β-氧化事件的其他产物,直到整个酰基辅酶A分子被还原为一组乙酰辅酶A分子。
脂肪酸以甘油三酯的形式储存在脂肪组织的脂肪库中。在两餐之间,它们的释放方式如下:
- 脂肪分解,去除从它们在它们的存储形式结合作为甘油三酯(或脂肪)的甘油的脂肪酸链,通过进行脂肪酶。这些脂肪酶被血液中高水平的肾上腺素和胰高血糖素(或由交感神经在脂肪组织中分泌的去甲肾上腺素)激活,由餐后血糖水平下降引起,同时降低血液中的胰岛素水平。
- 一旦从甘油中释放出来,游离脂肪酸就会进入血液,血液中的脂肪酸会附着在血浆白蛋白上,输送到全身各处。
- 长链游离脂肪酸进入细胞代谢(即最活细胞在体内除了红血细胞和神经元在中枢神经系统)通过特定的转运蛋白,如SLC27家族脂肪酸转运蛋白。红细胞不含线粒体,因此不能代谢脂肪酸;尽管含有线粒体,但中枢神经系统的组织不能使用脂肪酸,因为长链脂肪酸(与中链脂肪酸相反)不能穿过血脑屏障进入沐浴这些细胞的间质液中。
- 一旦进入细胞内,长链脂肪酸-CoA连接酶会催化脂肪酸分子与ATP(分解为AMP和无机焦磷酸盐)之间的反应,生成脂肪酰基腺苷酸,然后与游离辅酶A反应得到脂肪酰基辅酶A分子。
- 为了使酰基辅酶A进入线粒体,使用肉碱穿梭:
- 酰基辅酶A转移到肉碱的羟基通过肉碱棕榈我,位于的胞质面外和内线粒体膜。
- 酰基肉碱通过肉碱酰基肉碱移位酶在内部穿梭,就像肉碱穿梭在外面一样。
- 酰基肉碱被转换回酰基辅酶A通过肉碱棕榈II,位于内部面内线粒体膜。释放的肉碱穿梭回胞质溶胶,因为酰基辅酶A穿梭到线粒体基质中。
- β氧化,在线粒体基质中,然后将脂肪酸的长碳链(以酰基辅酶A分子的形式)切割成一系列双碳(醋酸盐)单元,这些单元与辅酶A结合形成乙酰辅酶A分子,在柠檬酸循环的“开始”与草酰乙酸缩合形成柠檬酸盐。可以方便地将此反应视为循环的“起点”,因为此时燃料-乙酰辅酶A-被添加到循环中,将作为CO2和H2消散在循环的每一轮过程中,释放大量以ATP形式捕获的能量。
简而言之,β氧化(游离脂肪酸最初分解为乙酰辅酶A)的步骤如下:
- 通过酰基辅酶A脱氢酶脱氢,产生1FADH2
- 水合通过烯酰辅酶A水
- 脱氢由3-羟酰辅酶A脱氢酶,得到1NADH+H+
- 被硫解酶切割,产生1个乙酰辅酶A和一个现在已经缩短了2个碳的脂肪酸(形成一个新的、缩短的酰基辅酶A)
重复这种β氧化反应,直到脂肪酸完全还原为乙酰辅酶A,或者在具有奇数个碳原子的脂肪酸的情况下,乙酰辅酶A和每分子脂肪酸1分子丙酰辅酶A。酰基辅酶A分子的每个β氧化切割产生5个ATP分子。
- β氧化产生的乙酰辅酶A通过与草酰乙酸结合形成柠檬酸进入线粒体中的柠檬酸循环。这导致乙酰辅酶A完全燃烧成CO2和水。在这个过程中释放的能量以每氧化一个乙酰辅酶A分子1个GTP和11个ATP分子的形式被捕获。这是乙酰辅酶A在脂肪酸β氧化发生的任何地方的命运,肝脏中的某些情况除外。
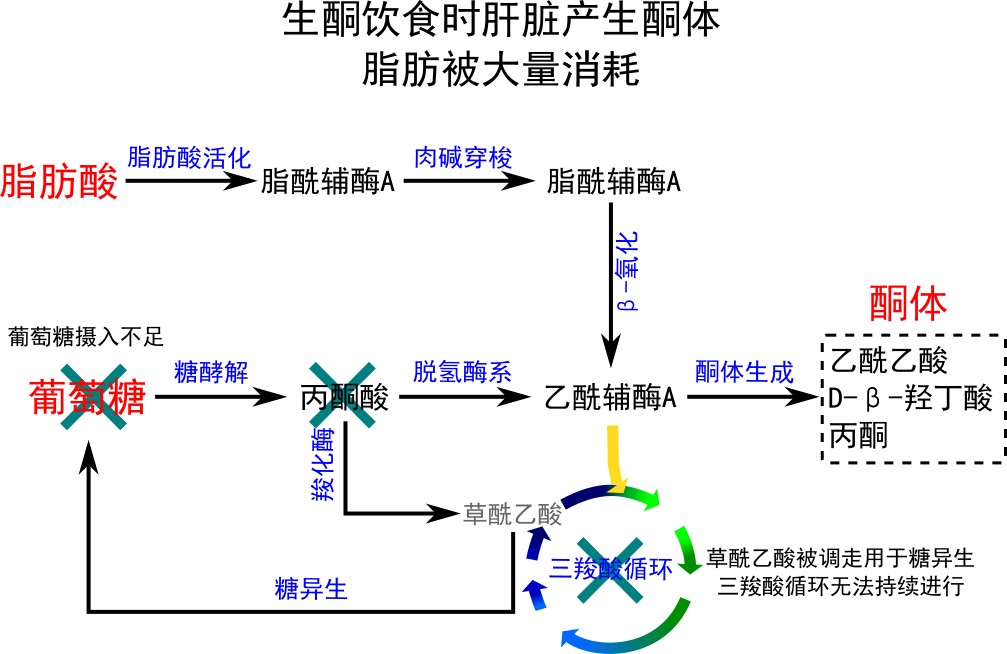
在禁食、饥饿、低碳水化合物饮食、长时间剧烈运动和不受控制的1型糖尿病期间,肝脏中的草酰乙酸可以全部或部分转移到糖异生途径中。在这些情况下,草酰乙酸被氢化成苹果酸,然后从肝细胞的线粒体中去除,在肝细胞的细胞质中转化为葡萄糖,从那里它被释放到血液中。因此,当低(或缺乏)胰岛素和高胰高血糖素刺激显着糖异生时,在肝脏中,草酰乙酸不能与乙酰辅酶A缩合血液中的浓度。在这些情况下,乙酰辅酶A转向形成乙酰乙酸盐和β-羟基丁酸盐。乙酰乙酸盐、β-羟基丁酸盐及其自发分解产物丙酮经常被称为酮体,但容易混淆(因为它们根本不是“物体”,而是水溶性化学物质)。酮体由肝脏释放到血液中。所有带有线粒体的细胞都可以从血液中吸收酮并将它们重新转化为乙酰辅酶A,然后可以将其用作柠檬酸循环中的燃料,因为没有其他组织可以将其草酰乙酸以这种方式转移到糖异生途径中发生在肝脏。与游离脂肪酸不同,酮可以穿过血脑屏障,因此可作为中枢神经系统细胞的燃料,作为葡萄糖的替代品,这些细胞通常靠葡萄糖生存。在饥饿、低碳水化合物饮食、长时间剧烈运动和不受控制的1型糖尿病期间,血液中出现高水平的酮被称为酮症,在其极端形式中,在失控的1型糖尿病中,如酮症酸中毒。
脂肪酶作用释放的甘油被肝脏(唯一可以发生这种反应的组织)中的甘油激酶磷酸化,生成的3-磷酸甘油被氧化成磷酸二羟丙酮。糖酵解酶磷酸丙糖异构酶将该化合物转化为3-磷酸甘油醛,后者通过糖酵解被氧化,或通过糖异生转化为葡萄糖。
脂肪酸作为能量来源
编辑脂肪酸以甘油三酯的形式储存在生物体中,是一种重要的能量来源,因为它们既是还原的又是无水的。一克脂肪酸的能量产量约为9kcal(37kJ),而碳水化合物则为4kcal(17kJ)。由于脂肪酸的烃部分是疏水的,这些分子可以储存在相对无水(无水)的环境中。另一方面,碳水化合物的水分含量更高。例如,1克糖原可以结合大约2克水,转换为1.33kcal/g(4kcal/3g)。这意味着脂肪酸每单位存储质量可以容纳六倍以上的能量。换句话说,如果人体依靠碳水化合物来储存能量,那么一个人需要携带31公斤(67.5磅)的水合糖原才能获得相当于4.6公斤(10磅)脂肪的能量。
冬眠动物为利用脂肪储备作为燃料提供了一个很好的例子。例如,熊冬眠约7个月,在这整个期间,能量来自脂肪储存的降解。候鸟在开始洲际旅行之前同样会积累大量的脂肪储备。
因此,年轻成人的脂肪储存量平均在10-20公斤之间,但因年龄、性别和个人性格而有很大差异。相比之下,人体仅储存约400克糖原,其中300克被锁定在骨骼肌内,不能被整个身体利用。储存在肝脏中的100克左右的糖原会在饥饿一天内耗尽。此后,由肝脏释放到血液中供身体组织普遍使用的葡萄糖必须由生糖氨基酸和其他一些不包括脂肪酸的糖异生底物合成。但是请注意,脂肪分解会释放甘油,甘油可以进入糖异生途径。
由甘油和脂肪酸合成碳水化合物
编辑脂肪酸通过线粒体内的β氧化分解为乙酰辅酶A,而脂肪酸由线粒体外的乙酰辅酶A在细胞质中合成。这两种途径是不同的,不仅在它们发生的位置上,而且在发生的反应和使用的底物上也是不同的。两条途径相互抑制,阻止β-氧化产生的乙酰辅酶A通过乙酰辅酶A羧化酶反应进入合成途径。它也不能转化为丙酮酸,因为丙酮酸脱氢酶复合反应是不可逆的。相反,脂肪酸β-氧化产生的乙酰辅酶A与草酰乙酸缩合,进入柠檬酸循环。在循环的每一轮中,在异柠檬酸脱氢酶和α-酮戊二酸脱氢酶催化的脱羧反应中,两个碳原子以CO2的形式离开循环。因此,柠檬酸循环的每一轮都会氧化乙酰辅酶A单元,同时再生最初与乙酰辅酶A结合形成柠檬酸的草酰乙酸分子。脱羧反应发生在循环中形成苹果酸之前。只有植物拥有将乙酰辅酶A转化为草酰乙酸的酶,草酰乙酸可以从中形成苹果酸,最终转化为葡萄糖。
然而,乙酰辅酶A可以转化为乙酰乙酸,乙酰乙酸可以脱羧为丙酮(自发或通过乙酰乙酸脱羧酶)。然后它可以进一步代谢为异丙醇,后者在呼吸/尿液中排泄,或通过CYP2E1代谢为羟基丙酮(丙酮醇)。丙酮醇可以转化为丙二醇。这会转化为甲酸和乙酸(后者转化为葡萄糖),或丙酮酸(通过两种替代酶),或丙醛,或先转化为L-乳醛,然后转化为L-乳酸盐(常见的乳酸异构体)。另一种途径将丙酮醇转化为甲基乙二醛,然后转化为丙酮酸,或转化为D-乳醛(通过SD-乳酰-谷胱甘肽或其他方式),然后转化为D-乳酸酯。D-乳酸代谢(转化为葡萄糖)在人体中缓慢或受损,因此大部分D-乳酸通过尿液排出;因此,源自丙酮的D-乳酸可显着导致与酮症或异丙醇中毒相关的代谢性酸中毒。升-乳酸可以完成脂肪酸向葡萄糖的净转化。1951年进行了第一个显示丙酮转化为葡萄糖的实验。这和进一步的实验使用碳同位素标记。高达11%的葡萄糖可以在人类饥饿期间从丙酮中提取。
脂肪组织中甘油三酯在脂解过程中释放到血液中的甘油只能被肝脏吸收。在这里,它通过甘油激酶的作用转化为甘油3-磷酸,甘油激酶水解每被磷酸化的甘油分子一分子ATP。然后将3-磷酸甘油氧化为磷酸二羟基丙酮,后者又通过磷酸丙糖异构酶转化为3-磷酸甘油醛。从这里开始,原始甘油的三个碳原子可以通过糖酵解被氧化,或通过糖异生转化为葡萄糖。
脂肪酸的其他功能和用途
编辑细胞内信号
编辑脂肪酸是构成大部分的磷脂的组成部分质膜,或细胞膜,细胞。这些磷脂可以通过细胞膜结合酶磷脂酶C(PLC)水解磷脂、磷脂酰肌醇4,5-二磷酸酯(PIP2)裂解为二酰基甘油(DAG)和三磷酸肌醇(IP3)。
类二十烷酸旁分泌激素
编辑脂肪酸代谢的一种产物是前列腺素,这种化合物在动物体内具有多种激素样作用。在人类和其他动物的几乎所有组织中都发现了前列腺素。它们通过酶促衍生自花生四烯酸,一种20碳多不饱和脂肪酸。因此,每个前列腺素含有20个碳原子,包括一个5碳环。它们是类二十烷酸的一个亚类,形成脂肪酸衍生物的前列腺素类。
前列腺素是通过从构成细胞膜的磷脂上裂解花生四烯酸而在细胞膜中合成的。这由直接作用于膜磷脂的磷脂酶A2或作用于DAG(二酰基甘油)的脂肪酶催化。然后花生四烯酸被前列腺素合酶的环氧合酶成分作用。这在脂肪酸链的大致中间形成环戊烷环。该反应还添加了源自两个O2分子的4个氧原子。产生的分子是前列腺素G2,它被酶复合物的氢过氧化物酶成分转化为前列腺素H2.这种高度不稳定的化合物会迅速转化为其他前列腺素、前列环素和血栓素。然后这些物质被释放到制造类二十烷酸激素的细胞周围的间质液中。
如果花生四烯酸被脂加氧酶而不是环加氧酶作用,则形成羟基二十碳四烯酸和白三烯。它们还充当局部激素。
前列腺素有两种衍生物:前列环素和血栓素。前列环素是强大的局部血管扩张剂,可抑制血小板聚集。通过它们在血管舒张中的作用,前列环素也参与炎症。它们在血管壁中合成,具有防止不必要的凝块形成以及调节平滑肌组织收缩的生理功能。相反,血栓素(由血小板细胞产生)是血管收缩剂,促进血小板聚集。它们的名字来自它们在凝块形成(血栓形成)中的作用)。
脂肪酸的膳食来源
编辑膳食脂肪在十二指肠中被肥皂以胆汁盐和磷脂(如磷脂酰胆碱)的形式乳化。这样形成的脂肪滴可以被胰脂肪酶攻击。胆汁酸(胆酸)的结构,以标准形式、半真实3D形式和图解3D形式表示在胆汁酸(例如胆酸)和脂肪、脂溶性维生素和胆固醇的消化产物存在下在十二指肠中形成的混合胶束的示意图。
人体中很大一部分脂肪酸以动物或植物来源的甘油三酯的形式从饮食中获得。从陆地动物获得的脂肪中的脂肪酸往往是饱和的,而鱼类和植物的甘油三酯中的脂肪酸通常是多不饱和的,因此以油的形式存在。
这些甘油三酯,不能被肠道吸收。它们被胰脂肪酶分解成甘油单酯和甘油二酯以及游离脂肪酸(但不含游离甘油),胰脂肪酶与称为辅脂肪酶的蛋白质(也是胰液的成分)形成1:1的复合物,是其活动所必需的。活化的复合物只能在水-脂肪界面起作用。因此,对于这些酶的最佳活性,必须首先通过胆汁盐乳化脂肪。由甘油三酯、甘油二酯和甘油单酯以及游离脂肪酸的混合物组成的消化产物,与饮食中的其他脂溶性成分(例如脂溶性维生素和胆固醇)和胆汁盐一起形成混合胶束,在水样十二指肠内容物(见右图)。
这些胶束的内容物(但不是胆汁盐)进入肠上皮细胞(小肠内壁的上皮细胞),在那里它们被重新合成为甘油三酯,并包装成乳糜微粒,然后释放到乳糜管(肠道淋巴系统的毛细血管)中)。这些乳汁排入胸导管它在颈部左下方的左颈静脉和左锁骨下静脉交界处排入静脉血。这意味着消化的脂溶性产物直接排入全身循环,而不像所有其他消化产物那样首先通过肝脏。这种特殊性的原因是未知的。
乳糜微粒在全身循环,使血浆在高脂肪餐后呈乳白色或奶油状。脂蛋白脂酶的上内皮表面毛细管的,特别是在脂肪组织,但在较小程度上也在其它组织中,部分消化乳糜微粒成游离脂肪酸,甘油和乳糜微粒残余物。脂肪酸被脂肪细胞吸收,但甘油和乳糜微粒会残留。留在血浆中,最终被肝脏从循环中清除。乳糜微粒消化释放的游离脂肪酸被脂肪细胞吸收,在那里它们使用从糖酵解途径中的葡萄糖衍生的甘油重新合成为甘油三酯。这些甘油三酯储存在脂肪细胞的脂肪滴中,直到需要其他组织的燃料需求为止。
在肝脏吸收来自血液中葡萄糖的比例门静脉从肠道的到来。在肝脏补充了它的糖原储备(满时仅约100克糖原)后,大部分剩余的葡萄糖将转化为脂肪酸,如下所述。这些脂肪酸与甘油结合形成甘油三酯,它们被包装成与乳糜微粒非常相似的液滴,但被称为极低密度脂蛋白(VLDL)。这些VLDL液滴的处理方式与乳糜微粒完全相同,只是VLDL残留物被称为中密度脂蛋白(IDL),能够清除血液中的胆固醇。这会将IDL转化为低密度脂蛋白(LDL),后者被需要胆固醇纳入其细胞膜或用于合成目的(例如类固醇激素的形成)的细胞吸收。其余的低密度脂蛋白被肝脏清除。
脂肪组织和哺乳期乳腺也会从血液中吸收葡萄糖以转化为甘油三酯。这与在肝脏中的发生方式相同,不同之处在于这些组织不会将由此产生的甘油三酯以VLDL的形式释放到血液中。当胰岛素的血浆浓度低而胰高血糖素和/或肾上腺素的血浆浓度高时,脂肪组织细胞将甘油三酯储存在它们的脂肪滴中,最终将它们作为游离脂肪酸和甘油再次释放到血液中.乳腺在垂体前叶激素催乳素的影响下将脂肪(作为奶油脂肪滴)排放到乳汁中。
身体中的所有细胞都需要制造和维持它们的膜和细胞器的膜。他们是否完全依赖从血液中吸收的游离脂肪酸,或者能够从血糖合成自己的脂肪酸,尚不清楚。中枢神经系统的细胞几乎肯定有能力制造自己的脂肪酸,因为这些分子不能通过血脑屏障到达它们,而另一方面,体内没有细胞可以制造所需的必需脂肪酸。必须从饮食中获取并通过血液输送到每个细胞的酸。
脂肪酸合成
编辑与β-氧化非常相似,直链脂肪酸的合成通过如下所示的六个重复反应进行,直到产生16个碳的棕榈酸。
所提供的图表显示了脂肪酸是如何在微生物中合成的,并列出了在大肠杆菌中发现的酶。这些反应是由脂肪酸合成酶II(FASII)进行的,它通常包含多种酶作为一种复合物。FASII存在于原核生物、植物、真菌和寄生虫以及线粒体中。
在动物以及酵母等一些真菌中,这些相同的反应也会发生在脂肪酸合酶I(FASI)上,FASI是一种大的二聚体蛋白质,具有生成脂肪酸所需的所有酶活性。FASI的效率低于FASII;然而,它允许通过早期链终止形成更多分子,包括“中链”脂肪酸。酶、酰基转移酶和转酰基酶通过在酰基受体和供体之间转移脂肪酸将脂肪酸结合到磷脂、三酰基甘油等中。它们还负责合成生物活性脂质及其前体分子。
疾病
编辑脂肪酸代谢障碍可以用例如高甘油三酯血症(甘油三酯水平过高)或其他类型的高脂血症来描述。这些可能是家族性的或获得性的。
脂肪酸代谢紊乱的家族性类型通常被归类为脂质代谢的先天性错误。这些疾病可被描述为脂肪酸氧化障碍或脂质储存障碍,并且是由于酶缺陷影响身体氧化脂肪酸以在肌肉内产生能量的能力而导致的几种先天性代谢错误中的任何一种,肝脏和其他细胞类型。
此外,癌细胞可以在脂肪酸合成和线粒体脂肪酸氧化(FAO)方面表现出不规则的脂肪酸代谢,这两个方面涉及肿瘤发生和细胞生长的各个方面。
内容由匿名用户提供,本内容不代表vibaike.com立场,内容投诉举报请联系vibaike.com客服。如若转载,请注明出处:https://vibaike.com/127833/